Уэллс, 1913
Страница 2 из 2
Страница 2 из 2 •  1, 2
1, 2
Re: Уэллс, 1913
автор Gudleifr Ср Май 19, 2021 12:25 am
ОТДЕЛ IV. ПОДЪЕМНЫЕ ЖЕЛЕЗНЫЕ ДОРОГИ, ГОРКИ ДЛЯ ШАРИКОВ, ЗАМКИ И ВОЕННЫЕ ИГРЫ,- НО ОЧЕНЬ НЕМНОГО ВОЕННЫХ ИГР.
Я дал ужо теперь два общих типа игр на полу; но в смысле удовольствия и работы воображения игры эти лишь два образца тех вариантов, которые могут быть выполнены из описанных мною игрушек. Сейчас я в кратких словах хочу коснуться и некоторых других случаев, когда прекрасно могут быть использованы пол, доски, бруски, фигурки, железнодорожный состав,- весь арсенал для успешного изгнания беса скуки из жизни маленьких детей. Тут на первом плане стоит забава, называемая нами подъемной железной дорогой, или фуникулером.

Временами острова почему-либо теряют для нас свою привлекательность, также и города кажутся уже чересчур методически-правильными, не доставляют много приключений, начинают стеснять нас, и нам хочется чего-нибудь нового - ну чего нибудь... шумного. Тогда мы говорим:
- Давайте строить фуникулер! Пусть у нас будет еще один, да такой, чтобы доходил до самого
стола!

И мы начинаем спорить,- может лучше построить горную железную дорогу? Уже одно название звучит приятно. Оно напоминает нам незабвенные времена, когда мы бродили в Венген, подымаясь по горам - из слякоти в такую светлую область снега и солнечного сияния, что глазам не верилось. И вот мы принимаемся делать горную железную дорогу.

Провести ее так далеко, до самого стола - это нам еще никогда не удавалось, но когда-нибудь удастся же! А потом мы решили иметь две станции - одну на столе, а другую на полу, и при каждой из них запасные и разъездные пути.

Привлекательная особенность горной железной дороги, раз она надлежащим образом сделана, заключается в том, что нагруженной платформа - а не игрушечный паровоз; слишком груба эта игра для хрупких деликатных паровозов - скатывается во весь опор с вершины горы вниз и летит по тому пути и так, как направят ваши искусно устроенные стрелки, а впоследствии - и это удивительное, но вполне понятное открытие,- вы можете пустить поезд обратно при помощи "лектрика".
Что такое "лектрик"? Это почти случайное открытие одного из нас, которому слово это обязано также своим происхождением. Явилось оно при одном приключении с игрушечным паровозом, казавшимся совершенно испорченным, когда на самом деле он был еще полон жизни.
Вы, вероятно, знаете, на что похож игрушечный паровоз? Общий вид его напоминает настоящий - те же трубы, буфера, корпус. Все эти вещи сделаны изящно, но это не способствует его легкости, не облегчает подъема в гору. Теперь дальше. Иногда в паровозе портится заводной механизм, и тогда он становится негодным; порою же портится только его наружный вид: труба согнута; корпус измят. Но вы снимаете эти части, и вот - у вас открытый заводной механизм на колесах,- аппарат почти неудержимой силы, точно дух без тела, что-то вроде неодушевленной ярости. Вот это-то младший член нашего семейства моментально окрестил словом "лектрик" и с тех пор стал трепать его...
(Кстати, я знал весьма пригодный маленький "лектрик", сделанный из заводной мыши).
Ну-с прекрасно! Когда у нас есть уже подпорки под рельсы - стулья, ящики и бруски - и мы хорошо и искусно уравняли нашу линию, облегчая тем спуск, тщательно соединили на поворотах рельсы, дабы спускающиеся платформы и повозки не соскочили с них,- мы посылаем вниз сначала пустую платформу, за нею платформы, нагруженные брусками и свинцовыми солдатиками, а затем уже то, что Уэллс-младший назвал "лектриком" (т.е. заводной паровоз); и вот сильный "лектрик" втаскивает платформы снова кверху с какой-то преднамеренной свирепостью и свистом, который чрезвычайно приятен нам. На линиях этих мы устроили стрелки; сделали мы на них же и горизонтальные разъезды, которые совершенно предотвращают катастрофы; линии эти шли вверх и вниз одна над другой, входили и выходили из туннелей...

Горка для шариков - большое сооружение, с которого мы делаем разветвленные спускающиеся пути, а по ним пускаем шарики [Игра в шарики очень распространена в Англии, заменяя там наши бабки. Шарики эти по большей части каменные, хотя встречаются глиняные и даже мраморные. Прим. перев.]. Не знаю, почему интересно заставлять катиться шарики по длинной извилистой дорожке, полуостанавливаясь, спускаться по ступенькам, устремляясь вдруг из скрытых мест и чрез маленькие картонные мосты, - но нам все это казалось интересным, и мы часто проделывали это.

Замки делаются из брусков и картонных башенок и оградами из тонких папок; тут же подъемный мост и ров с водой. Это просто особый род постройки городов, сделанный нами потому, что у нас был ящик с людьми в бронях. Мы могли бы воспроизвести всевозможные исторические эпохи, если бы фабриканты игрушечных солдатиков снабдили нас народом. Но в настоящее время,- о чем уже я имел случай пожалеть,- фабриканты почти не изготовляют ничего, кроме современных воинов. О военных же играх я должен был бы написать целые тома или ничего. На сей раз пусть это будет последнее... Когда-нибудь, быть может, я и возьмусь за пространную книгу о военных играх, буду говорить о сражениях и походах, о тактике и стратегии. Но сейчас я намеревался лишь рассказать об обычных радостях игры на полу и пробрать фабрикантов игрушек с целью усовершенствования и пользы дела. И насколько мне кажется,- мне это удалось. Если кому-либо из родителей или родственников удастся, благодаря мне, купить более умную игрушку, то жизнь моя не пройдет совсем напрасно.

Я дал ужо теперь два общих типа игр на полу; но в смысле удовольствия и работы воображения игры эти лишь два образца тех вариантов, которые могут быть выполнены из описанных мною игрушек. Сейчас я в кратких словах хочу коснуться и некоторых других случаев, когда прекрасно могут быть использованы пол, доски, бруски, фигурки, железнодорожный состав,- весь арсенал для успешного изгнания беса скуки из жизни маленьких детей. Тут на первом плане стоит забава, называемая нами подъемной железной дорогой, или фуникулером.

Временами острова почему-либо теряют для нас свою привлекательность, также и города кажутся уже чересчур методически-правильными, не доставляют много приключений, начинают стеснять нас, и нам хочется чего-нибудь нового - ну чего нибудь... шумного. Тогда мы говорим:
- Давайте строить фуникулер! Пусть у нас будет еще один, да такой, чтобы доходил до самого
стола!

И мы начинаем спорить,- может лучше построить горную железную дорогу? Уже одно название звучит приятно. Оно напоминает нам незабвенные времена, когда мы бродили в Венген, подымаясь по горам - из слякоти в такую светлую область снега и солнечного сияния, что глазам не верилось. И вот мы принимаемся делать горную железную дорогу.

Провести ее так далеко, до самого стола - это нам еще никогда не удавалось, но когда-нибудь удастся же! А потом мы решили иметь две станции - одну на столе, а другую на полу, и при каждой из них запасные и разъездные пути.

Привлекательная особенность горной железной дороги, раз она надлежащим образом сделана, заключается в том, что нагруженной платформа - а не игрушечный паровоз; слишком груба эта игра для хрупких деликатных паровозов - скатывается во весь опор с вершины горы вниз и летит по тому пути и так, как направят ваши искусно устроенные стрелки, а впоследствии - и это удивительное, но вполне понятное открытие,- вы можете пустить поезд обратно при помощи "лектрика".

Что такое "лектрик"? Это почти случайное открытие одного из нас, которому слово это обязано также своим происхождением. Явилось оно при одном приключении с игрушечным паровозом, казавшимся совершенно испорченным, когда на самом деле он был еще полон жизни.

Вы, вероятно, знаете, на что похож игрушечный паровоз? Общий вид его напоминает настоящий - те же трубы, буфера, корпус. Все эти вещи сделаны изящно, но это не способствует его легкости, не облегчает подъема в гору. Теперь дальше. Иногда в паровозе портится заводной механизм, и тогда он становится негодным; порою же портится только его наружный вид: труба согнута; корпус измят. Но вы снимаете эти части, и вот - у вас открытый заводной механизм на колесах,- аппарат почти неудержимой силы, точно дух без тела, что-то вроде неодушевленной ярости. Вот это-то младший член нашего семейства моментально окрестил словом "лектрик" и с тех пор стал трепать его...
(Кстати, я знал весьма пригодный маленький "лектрик", сделанный из заводной мыши).

Ну-с прекрасно! Когда у нас есть уже подпорки под рельсы - стулья, ящики и бруски - и мы хорошо и искусно уравняли нашу линию, облегчая тем спуск, тщательно соединили на поворотах рельсы, дабы спускающиеся платформы и повозки не соскочили с них,- мы посылаем вниз сначала пустую платформу, за нею платформы, нагруженные брусками и свинцовыми солдатиками, а затем уже то, что Уэллс-младший назвал "лектриком" (т.е. заводной паровоз); и вот сильный "лектрик" втаскивает платформы снова кверху с какой-то преднамеренной свирепостью и свистом, который чрезвычайно приятен нам. На линиях этих мы устроили стрелки; сделали мы на них же и горизонтальные разъезды, которые совершенно предотвращают катастрофы; линии эти шли вверх и вниз одна над другой, входили и выходили из туннелей...

Горка для шариков - большое сооружение, с которого мы делаем разветвленные спускающиеся пути, а по ним пускаем шарики [Игра в шарики очень распространена в Англии, заменяя там наши бабки. Шарики эти по большей части каменные, хотя встречаются глиняные и даже мраморные. Прим. перев.]. Не знаю, почему интересно заставлять катиться шарики по длинной извилистой дорожке, полуостанавливаясь, спускаться по ступенькам, устремляясь вдруг из скрытых мест и чрез маленькие картонные мосты, - но нам все это казалось интересным, и мы часто проделывали это.

Замки делаются из брусков и картонных башенок и оградами из тонких папок; тут же подъемный мост и ров с водой. Это просто особый род постройки городов, сделанный нами потому, что у нас был ящик с людьми в бронях. Мы могли бы воспроизвести всевозможные исторические эпохи, если бы фабриканты игрушечных солдатиков снабдили нас народом. Но в настоящее время,- о чем уже я имел случай пожалеть,- фабриканты почти не изготовляют ничего, кроме современных воинов. О военных же играх я должен был бы написать целые тома или ничего. На сей раз пусть это будет последнее... Когда-нибудь, быть может, я и возьмусь за пространную книгу о военных играх, буду говорить о сражениях и походах, о тактике и стратегии. Но сейчас я намеревался лишь рассказать об обычных радостях игры на полу и пробрать фабрикантов игрушек с целью усовершенствования и пользы дела. И насколько мне кажется,- мне это удалось. Если кому-либо из родителей или родственников удастся, благодаря мне, купить более умную игрушку, то жизнь моя не пройдет совсем напрасно.


Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Ср Май 19, 2021 12:33 am
Описание игры "Город", сильно напоминает довоенную книжку Бориса Житкова "Что я видел" -  PDF, 29.3Мб. А тогдашние понятия о жизни по нынешним временам считается еще той фантастикой. Что еще более делает СССР возможной для отыгрывания "волшебной страной". Правда, попутно возникает вопрос, что интереснее - играть или учиться. Особенно, если вспомнить еще более известное описание игры - "Швамбранию".
PDF, 29.3Мб. А тогдашние понятия о жизни по нынешним временам считается еще той фантастикой. Что еще более делает СССР возможной для отыгрывания "волшебной страной". Правда, попутно возникает вопрос, что интереснее - играть или учиться. Особенно, если вспомнить еще более известное описание игры - "Швамбранию".
Идея оживляжа "Островов" -ТЕМА #98, АБЗАЦ #1314.
... Можно и попытаться соединить "кубики" электрически. Например, на подставках солдатиков ТЕМА #91, АБЗАЦ #1168
ТЕМА #91, АБЗАЦ #1168

разместить узлы решетки
чтобы получить "машину боя"ТЕМА #69, АБЗАЦ #752

PDF, 29.3Мб. А тогдашние понятия о жизни по нынешним временам считается еще той фантастикой. Что еще более делает СССР возможной для отыгрывания "волшебной страной". Правда, попутно возникает вопрос, что интереснее - играть или учиться. Особенно, если вспомнить еще более известное описание игры - "Швамбранию".Идея оживляжа "Островов" -
ТЕМА #98, АБЗАЦ #1314.... Можно и попытаться соединить "кубики" электрически. Например, на подставках солдатиков
ТЕМА #91, АБЗАЦ #1168
разместить узлы решетки

чтобы получить "машину боя"
ТЕМА #69, АБЗАЦ #752
Последний раз редактировалось: Gudleifr (Вс Сен 08, 2024 2:24 am), всего редактировалось 4 раз(а)
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Чт Май 20, 2021 1:00 am
Вспомнил об этой статье, посетив Форум "современных игроделов". Вот пара перлов оттуда:
Т.е. компьютер - только хранилище картинок, а программист - лишь посредник в их переформатировании, ни о какой кибернетике речь не идет в принципе.
Вот и решил показать, как играли настоящие кибернетики.- G.
***
Jacobson H., On models of reproduction, American Scientist, 46, #3 (1958), 255-268.
Перевод И.В.Соловьева / Кибернетический сборник #7, 1963.
В течение последнего столетия ученым удалось построить неживые модели большинства характерных функций живых существ. Такие функции, как движение, энергетический обмен с внешней средой, накопление энергии, восприятие и реакции на внешние раздражители, выполнялись и прежде различными устройствами, например, соответственно автомобилем, паровой машиной, аккумуляторной батареей, иконоскопом и цифровой вычислительной машиной. Лишь одна основная функция, а именно воспроизведение (вытекающей из нее функцией можно считать рост) до сих пор не была удовлетворительно представлена неживой моделью. В данной статье описана конструкция простой самовоспроизводящей модели ("самовоспроизведение" и "воспроизведение" здесь означают одно и то же) из электромеханических элементов. В статье описывается также построение более полных моделей воспроизведения, разбирается сущность процесса воспроизведения как абстрактного принципа и необходимые условия для того, чтобы система была самовоспроизводящей. Исследуется аналогия между электромеханическими и биохимическими самовоспроизводящими системами. На основании изучения этих моделей сделаны выводы о возможности самопроизвольного зарождения жизни на земле.
I. ЭЛЕКТРОМЕХАНИЧЕСКИЕ АНАЛОГИ ВОСПРОИЗВЕДЕНИЯ
Воспроизведение является, по существу, столь простым процессом, что отсутствие до сего времени действующей неживой модели воспроизведения представляется удивительным. Определение процесса воспроизведения является до некоторой степени спорным. Мы отложим его до второго раздела. В настоящем разделе мы просто опишем задуманные, спроектированные и построенные модели.
ПРЕДВАРИТЕЛЬНЫЕ СООБРАЖЕНИЯ О ВОЗМОЖНЫХ МОДЕЛЯХ ВОСПРОИЗВЕДЕНИЯ
Очевидными неотъемлемыми факторами в любом процессе воспроизведения являются: 1) окружающая среда, в которой свободно перемещаются случайные элементы или части; 2) достаточное количество таких частей; 3) пригодный источник энергии для сборки этих частей и 4) случайно или намеренно собранный "протоиндивидуум", составленный из имеющихся частей и способный брать их из окружающей среды и синтезировать их в свою собственную функциональную копию, используя для этого имеющийся источник энергии.
Необходимые части делятся на различные классы; необходимы, по-видимому, следующие: 1) преобразователь энергии, превращающий энергию окружающей среды в отрицательную энтропию открытой системы, т.е. двигатель, использующий имеющуюся энергию для движения и упорядочения частей; 2) образ структуры, отражающий организацию индивидуума, который система может использовать для приведения в порядок имеющихся частей, т.е. план; 3) прибор для отбора (опознавания правильных и неправильных) частей в течение сборки, т.е. распознающая система.
Грубо мы можем сравнить эти три необходимых класса с эффекторами (например, мышцами), наследственными структурами (например, хромосомами) и воспринимающими органами (например, глазами) живых систем.
Первой механической моделью воспроизведения, пришедшей на ум автору и другим, занимавшимся этой проблемой, было простое механическое самособирающееся устройство, состоящее из двигателя, органа, хранящего план организации, и распознающего органа. Двигатель под управлением плана мог перемещать распознающий орган среди беспорядочно расположенных деталей, выбирать надлежащую деталь, перемещать ее на какой-нибудь соседний сборочный участок и выполнять необходимые сборочные операции. При соответствующем построении можно было даже монтировать детали. Однако основная трудность при построении такой модели состоит в том, чтобы спроектировать двигатель, который действовал бы на детали на расстоянии, выбирая нужную деталь и устанавливая ее на точно обусловленное место, находящееся на некотором расстоянии.
ДЕЙСТВУЮЩАЯ МОДЕЛЬ ВОСПРОИЗВЕДЕНИЯ
Указанное выше затруднение - создание двигателя для перемещения всех деталей - можно устранить, заставив детали двигаться под действием своей собственной энергии, т.е. сделав их самодвижущимися. Такую систему с самодвижущимися деталями можно легко построить. Это можно сделать следующим образом. Пусть детали представляют различного рода самодвижущиеся тележки. Окружающая среда - замкнутая дорожка, на которой тележки расположены в совершенно случайной последовательности, определенной какой-нибудь вероятностной процедурой, например бросанием жребия. "Организм" (кавычки всюду в этой статье указывают на модель соответствующих живых систем или процессов) представляет собой группу из двух или большего числа тележек, расположенных в заданном порядке, способную создавать другие "организмы" из окружающей среды. "Организм" собирается на боковых ветках, отходящих от дорожки "среды", где они могут "поджидать" и отбирать детали для новых "организмов". "Воспроизведение" состоит в том, что отбираются в надлежащем порядке тележки, которые повторяют первоначальный "организм". Отбор тележек для направления на данную ветку в процессе сборки состоит в надлежащем автоматическом открывании и закрывании стрелок на дорожке. Для этого особенно пригодна модель железной дороги. Мы построили такое устройство на модели железной дороги и назвали его воспроизводящее последовательное устройство один - ВПУ I.

Рис.1. Общее расположение элементов ВПУ I.
Общее расположение частей ВПУ I показано на рис.1. Достаточно иметь тележки двух видов, так как "мозг" можно поместить на тележку одного типа, которую мы назовем "головной" или тележкой A, а "мышцы" и "глаза" - на другую "хвостовую" тележку или тележку B. Конечно, и тележка A, и тележка B имеют каждая свой двигатель для движения туда, куда их направляют стрелки, или просто для движения по замкнутой дорожке.
Последовательность сборочных операций следующая:
1. Сначала все тележки расположены случайно и движутся по дорожке.
2. Под действием случая или воли конструктора или, всего вероятнее, действий оператора, нажимающего кнопки стрелок, тележка А и затем тележка В оказываются собранными как первый "организм" на первой ветке. Этот этап изображен на рис.1.
3. Тележка А этого "организма" передает "сообщение" тележке B: "подстереги, когда тележка А на дорожке "среды" приблизится ко второй ветке. Когда она подойдет, открой стрелку на эту ветку". Эти указания выполняются, и тележка А въезжает на вторую ветку.
4. Когда тележка A проходит через стрелку на вторую ветку, "мышцы" тележки B первого "организма" закрывают стрелку, так как иначе все остальные тележки последуют за ней на эту ветку. Затем тележка B "сообщает" тележке A, что стрелка закрыта.
5. "Мозг" на тележке А "думает" и затем дает тележке В следующую измененную инструкцию: "подстереги, когда тележка B на дорожке "среды" приблизится ко второй ветке. Когда она подойдет, открой стрелку на эту ветку". Тележка B опять "выполняет" указание, и теперь на вторую ветку въезжает другая тележка B и становится за тележкой A, прибывшей перед этим.
6. Тележка B первого "организма" опять закрывает стрелку на вторую ветку, когда тележка B прибыла на ветку. Этим предотвращается прибытие других тележек на эту ветку. Первой тележке A передается "сообщение" об этом действии.
7. "Мозг" первой тележки A опять "размышляет" и затем приказывает своему партнеру - тележке B: "не переводи больше стрелок". Этим заканчивается активная "жизнь" первого "организма", т.е. "жизнь" первого "поколения".
8. Но на второй ветке теперь находится новый "организм"; это - "второе поколение"; оно тождественно первому, но расположено на следующей ветке. "Организм" имеет те же свойства по отношению к прибывающим "поездам", как и первый "организм", и отбирает по порядку другие тележки A и B, собирая их на третьей ветке. Таким образом он продолжает выполнять план "размножения".
9. Всякий вновь образованный "организм" продолжает "размножаться" линейно, пока в "среде" не будут исчерпаны все детали или пока не окажется свободных веток, или пока где-нибудь при совершении цикла не произойдет ошибка, т.е. "мутация". Такое явление? как и у живых существ, обычно гибельно.
БОЛЕЕ ПОЛНЫЕ МОДЕЛИ ВОСПРОИЗВЕДЕНИЯ
Не говоря уже о неправдоподобности того, что "воспроизведение" ВПУ I может выйти из своих границ или даже заменить обычные механизмы воспроизведения, следует сделать некоторые замечания и оговорки. Прежде всего ВПУ I имеет линейный цикл "воспроизведения". Один "родитель", "отец" имеет одного "ребенка", этот "ребенок" тоже одного, на этот раз - "внука", и т.д. Это, очевидно, не похоже на то, что происходит в природе или в любой устойчиво существующей воспроизводящей системе. У простейших бесполых живых форм "родитель" производит два отпрыска, неразличимых в дальнейшем процессе. Чтобы вид сохранился, коэффициент размножения, т.е. отношение численности данного поколения к численности предыдущего должно быть больше единицы. В противном случае гибель особей до созревания не будет восполняться и вид исчезнет. В ВПУ I "коэффициент размножения" равен точно единице. Это означает, что любой несчастный случай (например, неправильное действие реле) приведет к гибели "вида". Но вообще можно построить размножающуюся модель ВПУ II (здесь мы не касаемся некоторых очень сложных проблем технического воплощения), в которой "организм" собирает два или большее число новых "организмов", действующих таким же образом.

Рис.2. Общее расположение элементов ВПУ II.
Для этого в модели данного вида нужно иметь лишь разветвляющиеся пути, как показано на рис.2, и много деталей наряду с более сложной последовательностью открывания и закрывания стрелок при удвоении числа тележек в каждом новом "поколении". На рис.2 стрелки указывают двух потомков каждого "организма". На плане можно видеть четыре "поколения". Второе отличие между ВПУ I и воспроизведением живых существ заключается в том, что у последних среда произвольна. Если одно из назначений пищи - доставить элементы для построения организма, то можно сказать, что точки разветвления изображают "рты" для поступления "пищи". Поэтому можно сказать, что среда ВПУ I имеет, так сказать, набор "ртов" перед сборкой "организма". "Организм", согласно этой точке зрения, просто управляет открыванием и закрыванием этих "ртов". С другой стороны, стрелки можно рассматривать как ниши в окружающей среде, подобно пещерам на берегу моря, в которых может происходить поглощение пищи. С этой точки зрения их можно рассматривать как элемент окружающей среды.
Приняв более строгую точку зрения и считая, что стрелки представляют собой "рты", можно построить "организм" другого рода. Ничто не препятствует тому, чтобы "организм" собирал не только "головы" и "хвосты", но и "рты". Такая модель, которую мы можем назвать ВПУ III, будет нести на себе боковую ветку и стрелку на каждой тележке А или "голове". В начале сборки она положит их на тележку впереди себя. После этого стрелкой будут управлять уже существующие "организмы", как и в случае ВПУ I. Опять-таки механические трудности чисто практического характера помешали нам создать ВПУ III. ВПУ III, очевидно, представлял бы простую замкнутую дорожку.
Третье очевидное отличие связано с произвольностью среды. Мы замечаем, что распределение деталей в ней случайно, но одномерно. Хотя биохимические системы в основном двумерны и трехмерны, одномерная модель может вполне служить для упрощенного изображения необходимого случайного расположения частей.
Мы замечаем также, что в окружающей среде имеются некоторые вспомогательные приспособления: распорки, упоры боковых веток, выключатели двигателей. Их также можно рассматривать как элементы окружающей среды, необходимые для осуществления процесса, проводимого по основным правилам определения, рассмотренного во втором разделе статьи.
Более существенно четвертое отличие. Процесс воспроизведения можно разделить на две общие ступени: 1) дупликация "организма" согласно набору планов, заключенных в наследственных структурах его "родителей"; 2) дупликация и повторение планов.
Можно показать, что эти функции можно разделить на примере двух устройств, выполняющих соответственно первую и вторую ступени. Первое устройство - ВПУ I, а второе - печатная машина, печатающая планы для самой себя. Планы для ВПУ I очень просты, но они введены заранее в каждую тележку A, в виде пробивок в "хромосомах". Следовательно, ВПУ I воспроизводит не всего себя, но повторяет себя из частей, которые включают его планы. Наоборот, печатная машина производит планы, но не новые печатные машины.
Полная самовоспроизводящаяся модель должна выполнять обе задачи, т.е. собирать организмы по своим планам и производить также набор планов. Это также возможно, но, очевидно, гораздо труднее. Мы спроектировали, но не построили такое устройство - ВПУ IV.
Такая система должна иметь некоторый размножитель планов. Поскольку план есть "хромосома", строение которой напоминает перфокарту, через отверстия которой производится соединение контактов, размножитель планов должен, естественно, представлять собой некоторое перфорирующее устройство. Он будет пробивать отверстия в чистой карте. Каждый раз, когда "организм" собирает новый "организм", он должен также пробить новую карту-"хромосому", являющуюся копией исходной. Необходимо, чтобы этот "организм" передавал информацию следующему поколению, либо доставляя ему набор законченных штампов, либо передавая новому организму надлежащие сигналы, чтобы он мог заперфорировать свои собственные планы.

Рис.3. Общее расположение элементов ВПУ IV.
На рис.3 показана схема расположения ВПУ IV. Функции дупликации "организма" для следующего поколения и создания нового набора планов в ВПУ IV выполняются одновременно. "Организм" ВПУ IV состоит из трех частей, обозначенных A, B и C. Части B и C одинаковы за исключением того, что часть B содержит планы, а C содержит лишь чистую карту, на которой можно пробить новые планы. На схеме видны несколько тележек A и C и лишь одна тележка В. Если при возникновении "вида" на первой ветке находится один "организм", как на рис.3, а остальные тележки А и С случайно перемещаются в окружающей среде, может начаться воспроизведение самих "организмов" и планов.
Воспроизведение "организма" происходит путем распознавания и переключения соответствующих тележек на следующую боковую ветку, как описано для ВПУ I. Но поскольку других тележек В нет, ее нужно сделать из тележки С, прежде чем завершится воспроизведение. В ВПУ IV пустая "хромосомная" карта "родительского организма" пробивается и затем передается от тележки B) где она возникла, тележке С, которая этим превращается в другую тележку В. Затем новая тележка В выпускается в окружающую среду через показанную на рисунке обратную ветку. После этого из имеющихся тележек A и C и из тележки B можно собрать новый "организм".
Описанная операция напоминает процесс создания в реальном организме генетических структур, которые затем включаются в тело организма нового поколения. Здесь они становятся составной частью нового организма, а также моделью для дальнейшего построения генетических структур.
Пятое отличие ВПУ I и истинного воспроизведения состоит в том, что почти все реальные организмы, дифференцированные настолько, что у них имеются специальные половые клетки, воспроизводятся путем полового размножения, а на этот процесс ни одна из серии моделей ВПУ не рассчитана. Поскольку процесс полового размножения, как известно, играет основную роль в ускорении эволюции и поскольку все "мутации" в моделях ВПУ, наблюдавшиеся до сих пор, оказывались летальными, по-видимому, не имеет особого смысла проектировать модель ВПУ, повторяющую эту функцию высших организмов.
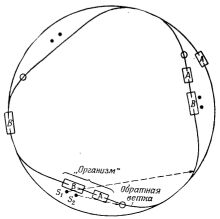
Рис.4. Общее расположение элементов ВПУ V.
Шестое отличие связано с окончательной судьбой частей, из которых составлены живые организмы. После смерти организма его части почти всегда используются снова другими организмами. В частности, животные используют довольно сложные остатки других организмов. Можно сделать довольно простую модель ВПУ V с повторным использованием частей. Нужно лишь предусмотреть возвращение тележек, образующих "организм", в окружающую среду после "смерти" "организма", т.е. после завершения цикла сборки. Это производится при помощи возвратных веток, изображенных на рис.4. Повторная сборка происходит тогда, когда "ребенок" берет части "родителя" для сборки "внука" (через две развилки дальше на дорожке).
Очевидно, при некотором усложнении схемы можно создать одну великолепную модель, сочетающую в себе все эти функции или любую их комбинацию.
- Т.е. "современный программист" якобы должен представить все тупо в уме, написать код, рассчитав (допустим он кодит платформер) всю геометрию уровней, размеры/скорости, противников, и весь интерактив встречающийся по ходу, ну и так же мысленно представляя расположение интерфейсов, их удобство и юзабилити полагаясь лишь на свой третий глаз? Конечно же разукрасив в коде все красивыми комментариями, чтоб представлять было удобней.
- А давайте в игры так же играть - поставим ведьмака на паузу, откинемся на кресле, закроем глаза и мысленно пройдем игру самым интересным для нас способом - а то современные игроки тоже обленились - все им во всех красках покажи да расскажи, за ручку проведи, в конце в попу поцелуй - совершенно не осталось места для фантазии и свободы действий.
- Большая часть проблем решается визуальными компонентами. Мне обычно хватает пару часов на создание всего этого. (Если без доп функций, типо выводов графиков, рассчетов, обработки данных и тп).
Т.е. компьютер - только хранилище картинок, а программист - лишь посредник в их переформатировании, ни о какой кибернетике речь не идет в принципе.
Вот и решил показать, как играли настоящие кибернетики.- G.
***
Jacobson H., On models of reproduction, American Scientist, 46, #3 (1958), 255-268.
Перевод И.В.Соловьева / Кибернетический сборник #7, 1963.
В течение последнего столетия ученым удалось построить неживые модели большинства характерных функций живых существ. Такие функции, как движение, энергетический обмен с внешней средой, накопление энергии, восприятие и реакции на внешние раздражители, выполнялись и прежде различными устройствами, например, соответственно автомобилем, паровой машиной, аккумуляторной батареей, иконоскопом и цифровой вычислительной машиной. Лишь одна основная функция, а именно воспроизведение (вытекающей из нее функцией можно считать рост) до сих пор не была удовлетворительно представлена неживой моделью. В данной статье описана конструкция простой самовоспроизводящей модели ("самовоспроизведение" и "воспроизведение" здесь означают одно и то же) из электромеханических элементов. В статье описывается также построение более полных моделей воспроизведения, разбирается сущность процесса воспроизведения как абстрактного принципа и необходимые условия для того, чтобы система была самовоспроизводящей. Исследуется аналогия между электромеханическими и биохимическими самовоспроизводящими системами. На основании изучения этих моделей сделаны выводы о возможности самопроизвольного зарождения жизни на земле.
I. ЭЛЕКТРОМЕХАНИЧЕСКИЕ АНАЛОГИ ВОСПРОИЗВЕДЕНИЯ
Воспроизведение является, по существу, столь простым процессом, что отсутствие до сего времени действующей неживой модели воспроизведения представляется удивительным. Определение процесса воспроизведения является до некоторой степени спорным. Мы отложим его до второго раздела. В настоящем разделе мы просто опишем задуманные, спроектированные и построенные модели.
ПРЕДВАРИТЕЛЬНЫЕ СООБРАЖЕНИЯ О ВОЗМОЖНЫХ МОДЕЛЯХ ВОСПРОИЗВЕДЕНИЯ
Очевидными неотъемлемыми факторами в любом процессе воспроизведения являются: 1) окружающая среда, в которой свободно перемещаются случайные элементы или части; 2) достаточное количество таких частей; 3) пригодный источник энергии для сборки этих частей и 4) случайно или намеренно собранный "протоиндивидуум", составленный из имеющихся частей и способный брать их из окружающей среды и синтезировать их в свою собственную функциональную копию, используя для этого имеющийся источник энергии.
Необходимые части делятся на различные классы; необходимы, по-видимому, следующие: 1) преобразователь энергии, превращающий энергию окружающей среды в отрицательную энтропию открытой системы, т.е. двигатель, использующий имеющуюся энергию для движения и упорядочения частей; 2) образ структуры, отражающий организацию индивидуума, который система может использовать для приведения в порядок имеющихся частей, т.е. план; 3) прибор для отбора (опознавания правильных и неправильных) частей в течение сборки, т.е. распознающая система.
Грубо мы можем сравнить эти три необходимых класса с эффекторами (например, мышцами), наследственными структурами (например, хромосомами) и воспринимающими органами (например, глазами) живых систем.
Первой механической моделью воспроизведения, пришедшей на ум автору и другим, занимавшимся этой проблемой, было простое механическое самособирающееся устройство, состоящее из двигателя, органа, хранящего план организации, и распознающего органа. Двигатель под управлением плана мог перемещать распознающий орган среди беспорядочно расположенных деталей, выбирать надлежащую деталь, перемещать ее на какой-нибудь соседний сборочный участок и выполнять необходимые сборочные операции. При соответствующем построении можно было даже монтировать детали. Однако основная трудность при построении такой модели состоит в том, чтобы спроектировать двигатель, который действовал бы на детали на расстоянии, выбирая нужную деталь и устанавливая ее на точно обусловленное место, находящееся на некотором расстоянии.
ДЕЙСТВУЮЩАЯ МОДЕЛЬ ВОСПРОИЗВЕДЕНИЯ
Указанное выше затруднение - создание двигателя для перемещения всех деталей - можно устранить, заставив детали двигаться под действием своей собственной энергии, т.е. сделав их самодвижущимися. Такую систему с самодвижущимися деталями можно легко построить. Это можно сделать следующим образом. Пусть детали представляют различного рода самодвижущиеся тележки. Окружающая среда - замкнутая дорожка, на которой тележки расположены в совершенно случайной последовательности, определенной какой-нибудь вероятностной процедурой, например бросанием жребия. "Организм" (кавычки всюду в этой статье указывают на модель соответствующих живых систем или процессов) представляет собой группу из двух или большего числа тележек, расположенных в заданном порядке, способную создавать другие "организмы" из окружающей среды. "Организм" собирается на боковых ветках, отходящих от дорожки "среды", где они могут "поджидать" и отбирать детали для новых "организмов". "Воспроизведение" состоит в том, что отбираются в надлежащем порядке тележки, которые повторяют первоначальный "организм". Отбор тележек для направления на данную ветку в процессе сборки состоит в надлежащем автоматическом открывании и закрывании стрелок на дорожке. Для этого особенно пригодна модель железной дороги. Мы построили такое устройство на модели железной дороги и назвали его воспроизводящее последовательное устройство один - ВПУ I.

Рис.1. Общее расположение элементов ВПУ I.
Общее расположение частей ВПУ I показано на рис.1. Достаточно иметь тележки двух видов, так как "мозг" можно поместить на тележку одного типа, которую мы назовем "головной" или тележкой A, а "мышцы" и "глаза" - на другую "хвостовую" тележку или тележку B. Конечно, и тележка A, и тележка B имеют каждая свой двигатель для движения туда, куда их направляют стрелки, или просто для движения по замкнутой дорожке.
Последовательность сборочных операций следующая:
1. Сначала все тележки расположены случайно и движутся по дорожке.
2. Под действием случая или воли конструктора или, всего вероятнее, действий оператора, нажимающего кнопки стрелок, тележка А и затем тележка В оказываются собранными как первый "организм" на первой ветке. Этот этап изображен на рис.1.
3. Тележка А этого "организма" передает "сообщение" тележке B: "подстереги, когда тележка А на дорожке "среды" приблизится ко второй ветке. Когда она подойдет, открой стрелку на эту ветку". Эти указания выполняются, и тележка А въезжает на вторую ветку.
4. Когда тележка A проходит через стрелку на вторую ветку, "мышцы" тележки B первого "организма" закрывают стрелку, так как иначе все остальные тележки последуют за ней на эту ветку. Затем тележка B "сообщает" тележке A, что стрелка закрыта.
5. "Мозг" на тележке А "думает" и затем дает тележке В следующую измененную инструкцию: "подстереги, когда тележка B на дорожке "среды" приблизится ко второй ветке. Когда она подойдет, открой стрелку на эту ветку". Тележка B опять "выполняет" указание, и теперь на вторую ветку въезжает другая тележка B и становится за тележкой A, прибывшей перед этим.
6. Тележка B первого "организма" опять закрывает стрелку на вторую ветку, когда тележка B прибыла на ветку. Этим предотвращается прибытие других тележек на эту ветку. Первой тележке A передается "сообщение" об этом действии.
7. "Мозг" первой тележки A опять "размышляет" и затем приказывает своему партнеру - тележке B: "не переводи больше стрелок". Этим заканчивается активная "жизнь" первого "организма", т.е. "жизнь" первого "поколения".
8. Но на второй ветке теперь находится новый "организм"; это - "второе поколение"; оно тождественно первому, но расположено на следующей ветке. "Организм" имеет те же свойства по отношению к прибывающим "поездам", как и первый "организм", и отбирает по порядку другие тележки A и B, собирая их на третьей ветке. Таким образом он продолжает выполнять план "размножения".
9. Всякий вновь образованный "организм" продолжает "размножаться" линейно, пока в "среде" не будут исчерпаны все детали или пока не окажется свободных веток, или пока где-нибудь при совершении цикла не произойдет ошибка, т.е. "мутация". Такое явление? как и у живых существ, обычно гибельно.
БОЛЕЕ ПОЛНЫЕ МОДЕЛИ ВОСПРОИЗВЕДЕНИЯ
Не говоря уже о неправдоподобности того, что "воспроизведение" ВПУ I может выйти из своих границ или даже заменить обычные механизмы воспроизведения, следует сделать некоторые замечания и оговорки. Прежде всего ВПУ I имеет линейный цикл "воспроизведения". Один "родитель", "отец" имеет одного "ребенка", этот "ребенок" тоже одного, на этот раз - "внука", и т.д. Это, очевидно, не похоже на то, что происходит в природе или в любой устойчиво существующей воспроизводящей системе. У простейших бесполых живых форм "родитель" производит два отпрыска, неразличимых в дальнейшем процессе. Чтобы вид сохранился, коэффициент размножения, т.е. отношение численности данного поколения к численности предыдущего должно быть больше единицы. В противном случае гибель особей до созревания не будет восполняться и вид исчезнет. В ВПУ I "коэффициент размножения" равен точно единице. Это означает, что любой несчастный случай (например, неправильное действие реле) приведет к гибели "вида". Но вообще можно построить размножающуюся модель ВПУ II (здесь мы не касаемся некоторых очень сложных проблем технического воплощения), в которой "организм" собирает два или большее число новых "организмов", действующих таким же образом.

Рис.2. Общее расположение элементов ВПУ II.
Для этого в модели данного вида нужно иметь лишь разветвляющиеся пути, как показано на рис.2, и много деталей наряду с более сложной последовательностью открывания и закрывания стрелок при удвоении числа тележек в каждом новом "поколении". На рис.2 стрелки указывают двух потомков каждого "организма". На плане можно видеть четыре "поколения". Второе отличие между ВПУ I и воспроизведением живых существ заключается в том, что у последних среда произвольна. Если одно из назначений пищи - доставить элементы для построения организма, то можно сказать, что точки разветвления изображают "рты" для поступления "пищи". Поэтому можно сказать, что среда ВПУ I имеет, так сказать, набор "ртов" перед сборкой "организма". "Организм", согласно этой точке зрения, просто управляет открыванием и закрыванием этих "ртов". С другой стороны, стрелки можно рассматривать как ниши в окружающей среде, подобно пещерам на берегу моря, в которых может происходить поглощение пищи. С этой точки зрения их можно рассматривать как элемент окружающей среды.
Приняв более строгую точку зрения и считая, что стрелки представляют собой "рты", можно построить "организм" другого рода. Ничто не препятствует тому, чтобы "организм" собирал не только "головы" и "хвосты", но и "рты". Такая модель, которую мы можем назвать ВПУ III, будет нести на себе боковую ветку и стрелку на каждой тележке А или "голове". В начале сборки она положит их на тележку впереди себя. После этого стрелкой будут управлять уже существующие "организмы", как и в случае ВПУ I. Опять-таки механические трудности чисто практического характера помешали нам создать ВПУ III. ВПУ III, очевидно, представлял бы простую замкнутую дорожку.
Третье очевидное отличие связано с произвольностью среды. Мы замечаем, что распределение деталей в ней случайно, но одномерно. Хотя биохимические системы в основном двумерны и трехмерны, одномерная модель может вполне служить для упрощенного изображения необходимого случайного расположения частей.
Мы замечаем также, что в окружающей среде имеются некоторые вспомогательные приспособления: распорки, упоры боковых веток, выключатели двигателей. Их также можно рассматривать как элементы окружающей среды, необходимые для осуществления процесса, проводимого по основным правилам определения, рассмотренного во втором разделе статьи.
Более существенно четвертое отличие. Процесс воспроизведения можно разделить на две общие ступени: 1) дупликация "организма" согласно набору планов, заключенных в наследственных структурах его "родителей"; 2) дупликация и повторение планов.
Можно показать, что эти функции можно разделить на примере двух устройств, выполняющих соответственно первую и вторую ступени. Первое устройство - ВПУ I, а второе - печатная машина, печатающая планы для самой себя. Планы для ВПУ I очень просты, но они введены заранее в каждую тележку A, в виде пробивок в "хромосомах". Следовательно, ВПУ I воспроизводит не всего себя, но повторяет себя из частей, которые включают его планы. Наоборот, печатная машина производит планы, но не новые печатные машины.
Полная самовоспроизводящаяся модель должна выполнять обе задачи, т.е. собирать организмы по своим планам и производить также набор планов. Это также возможно, но, очевидно, гораздо труднее. Мы спроектировали, но не построили такое устройство - ВПУ IV.
Такая система должна иметь некоторый размножитель планов. Поскольку план есть "хромосома", строение которой напоминает перфокарту, через отверстия которой производится соединение контактов, размножитель планов должен, естественно, представлять собой некоторое перфорирующее устройство. Он будет пробивать отверстия в чистой карте. Каждый раз, когда "организм" собирает новый "организм", он должен также пробить новую карту-"хромосому", являющуюся копией исходной. Необходимо, чтобы этот "организм" передавал информацию следующему поколению, либо доставляя ему набор законченных штампов, либо передавая новому организму надлежащие сигналы, чтобы он мог заперфорировать свои собственные планы.

Рис.3. Общее расположение элементов ВПУ IV.
На рис.3 показана схема расположения ВПУ IV. Функции дупликации "организма" для следующего поколения и создания нового набора планов в ВПУ IV выполняются одновременно. "Организм" ВПУ IV состоит из трех частей, обозначенных A, B и C. Части B и C одинаковы за исключением того, что часть B содержит планы, а C содержит лишь чистую карту, на которой можно пробить новые планы. На схеме видны несколько тележек A и C и лишь одна тележка В. Если при возникновении "вида" на первой ветке находится один "организм", как на рис.3, а остальные тележки А и С случайно перемещаются в окружающей среде, может начаться воспроизведение самих "организмов" и планов.
Воспроизведение "организма" происходит путем распознавания и переключения соответствующих тележек на следующую боковую ветку, как описано для ВПУ I. Но поскольку других тележек В нет, ее нужно сделать из тележки С, прежде чем завершится воспроизведение. В ВПУ IV пустая "хромосомная" карта "родительского организма" пробивается и затем передается от тележки B) где она возникла, тележке С, которая этим превращается в другую тележку В. Затем новая тележка В выпускается в окружающую среду через показанную на рисунке обратную ветку. После этого из имеющихся тележек A и C и из тележки B можно собрать новый "организм".
Описанная операция напоминает процесс создания в реальном организме генетических структур, которые затем включаются в тело организма нового поколения. Здесь они становятся составной частью нового организма, а также моделью для дальнейшего построения генетических структур.
Пятое отличие ВПУ I и истинного воспроизведения состоит в том, что почти все реальные организмы, дифференцированные настолько, что у них имеются специальные половые клетки, воспроизводятся путем полового размножения, а на этот процесс ни одна из серии моделей ВПУ не рассчитана. Поскольку процесс полового размножения, как известно, играет основную роль в ускорении эволюции и поскольку все "мутации" в моделях ВПУ, наблюдавшиеся до сих пор, оказывались летальными, по-видимому, не имеет особого смысла проектировать модель ВПУ, повторяющую эту функцию высших организмов.

Рис.4. Общее расположение элементов ВПУ V.
Шестое отличие связано с окончательной судьбой частей, из которых составлены живые организмы. После смерти организма его части почти всегда используются снова другими организмами. В частности, животные используют довольно сложные остатки других организмов. Можно сделать довольно простую модель ВПУ V с повторным использованием частей. Нужно лишь предусмотреть возвращение тележек, образующих "организм", в окружающую среду после "смерти" "организма", т.е. после завершения цикла сборки. Это производится при помощи возвратных веток, изображенных на рис.4. Повторная сборка происходит тогда, когда "ребенок" берет части "родителя" для сборки "внука" (через две развилки дальше на дорожке).
Очевидно, при некотором усложнении схемы можно создать одну великолепную модель, сочетающую в себе все эти функции или любую их комбинацию.
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Чт Май 20, 2021 1:25 am
II. ОБЩИЕ РАССУЖДЕНИЯ
ОПРЕДЕЛЕНИЕ ПОНЯТИЯ ВОСПРОИЗВЕДЕНИЯ
Мы описали пять очевидных моделей процесса воспроизведения, не определив сам процесс. При проектировании моделей было принято следующее неявное определение: воспроизведение есть действие упорядоченной индивидуальной структуры, которая, входя в некоторую систему, выбирает из окружающей среды случайно расположенные части и строит из них и из своих собственных частей функционально-эквивалентные структуры. Серия моделей ВПУ, очевидно, соответствует этому определению и, по существу, подсказывает его. Это определение исключает процессы, часто называемые процессами воспроизведения, но по интуитивному восприятию не являющиеся таковыми. Например, действия печатной машины или штамповальной машины не подходят под это определение, если только они не производят новые печатные или штамповальные машины.
При проектировании модели воспроизведения необходимо избегать двух опасностей: во-первых, опасности создать тривиально простую машину и, во-вторых, опасности создать нереализуемо сложную систему.
Для ВПУ были выбраны два рода деталей. Это, очевидно, наименьшее число, которое может быть введено в окружающую среду случайно. Число родов деталей можно свести к одному, но тогда не будет выполнено требование случайности. ВПУ I при помощи любопытного процесса можно свести к одноэлементной системе. При этом тележки А ставят на место, по одной на ветке, и устанавливают так, что их "хромосомы" находятся во второй позиции, т.е. их инструкции позволяют входить на ветку только тележке В. Если одна тележка В впускается на первую ветку, последующие ветки заполняются одна за другой тележками В. Хотя это прекрасный пример автоматического последовательного устройства, это уже не будет представлять воспроизведения.
Такой процесс напоминает кристаллизацию, которую также часто приводят как пример воспроизведения. Несомненно, при кристаллизации происходит усложнение структуры, однако в нем нет отчетливой индивидуализации. Ее нет даже в такой двухкомпонентной кристаллизующейся системе, как хлористый натрий. О таком кристалле никак нельзя сказать, что данный ион натрия или хлора произошел от какого-нибудь другого конкретного иона или данной группы ионов.
Часто в качестве аналога процесса воспроизведения приводят другую химическую параллель - автокатализ. Автокатализ также отличается от воспроизведения отсутствием индивидуальности. Для объяснения возникновения жизни, которое часто связывают с процессом кристаллизации или автокатализа, требуется большее, а именно объяснить происхождение индивидуальности, которая выражается в сходстве между родителями и потомками, наблюдаемом во всей живой природе. Некоторые виды кристаллизации и автокатализа были, без сомнения, предвестниками жизни, однако они не удовлетворяют нашему определению воспроизведения.
Определение неполно и в том отношении, что оно не указывает характер окружающей среды, допускающей воспроизведение. По-видимому, она может быть произвольной. Две абстрактные модели воспроизведения были упомянуты фон Нейманом [1, 2], который первый обосновал возможность создания подобных автоматов. В этих моделях в качестве первого типа окружающей среды была указана внутренняя структура перфорационной машины Тьюринга - мало вероятный случай среды; второй тип среды - своеобразная группа смежных прямоугольных клеток, причем изменения в одной клетке могут воздействовать на конечное число состояний в соседней клетке. Оба эти типа среды весьма произвольны, как и в случае моделей ВПУ. Единственное, что делает модели ВПУ шагом вперед по сравнению с системами фон Неймана - то, что они спроектированы в деталях, и по крайней мере одна из них работает.
Неприменимость таких произвольных типов среды заключается в их искусственности. Биологи ищут систему, действующую в среде, не требующей вмешательства человека, т.е. модель самопроизвольного возникновения самовоспроизводящегося вида. Жизнь, конечно, появилась не на рельсовом пути макета железной дороги! Но наши тележки следует принимать как модели, независимо от типа среды. По-видимому, их можно расценивать по изяществу, которое будет больше в моделях с более простым окружением, с меньшей аппаратурой и более простыми частями.
Мы осмеливаемся теперь высказать мысль, что этот процесс самоупорядочения можно ввести в электронную систему. Хотя в ней не будет наглядности движущихся частей и игрушечных поездов, устройство из триггерных схем, магнитных элементов или реле может быть сделано столь же близким к нашему определению, как любые из упомянутых моделей. Если построить системы электрических цепей таким образом, что упорядочение одной известным образом заставляет другие упорядочиваться таким же образом, то мы получим электронную модель воспроизведения.
АНАЛОГИ БИОЛОГИЧЕСКИХ СИСТЕМ И ИХ ФУНКЦИЙ
Ценность моделей этого рода состоит в том, что они привлекают наше внимание к абстрактным функциям, присущим моделируемым ими процессам. Модель, как и живое существо, имеет некоторый необходимый минимальный набор функций. Мы уже указали на абсолютную необходимость: а) источника составных частей, двигателя, планов и некоторого механизма опознавания, б) источника энергии и в) системы передачи и обработки информации или последовательного упорядочения. Их можно считать минимальными функциями, самовоспроизводящейся системы. Рассмотрим аналоги каждой из них.
а) СОСТАВНЫЕ ЧАСТИ. В состав живых существ входят вода, соли, аминокислоты, жирные кислоты и витамины для животных; вода, углекислота и соли для растений. Из этих простых частей организмы создают свои копии.
Из сравнения с моделями ВПУ видно, что живые существа представляют собой сложные собрания простых частей, а модели - простые собрания сложных частей. В этом смысле модели не представляют собой подлинные копии живых существ. Представляется удивительным, сколь малым числом составных частей может обойтись простое живое существо, обладая при этом способностью функционировать и воспроизводить самого себя. Это, конечно, зависит от того, что называть "частью". Если исходить из частей, уже имеющихся в готовом виде в окружающей среде, то моделям нужны две или три такие части, а живым существам - миллионы частей. Естественная, реальная, т.е. неискусственная среда, вряд ли содержит значительное число столь сложных частей, как описанные тележки A и B. Вероятность появления таких частей зависит от их сложности, от того, сколь часто их составные части встречаются в окружающей среде, и от свободной энергии устройства. Мы обошли это затруднение, изготовив части заранее. В природе устройства, равные по сложности тележкам A и B, могут не появиться в течение значительного времени. Поэтому вопрос о том, были ли первые живые существа собраниями небольшого числа сложных частей или большого числа простых частей, остается совершенно нерешенным. Всякая попытка сделать выводы по этому пункту может дать лишь весьма убогие результаты.
б) ЭНЕРГИЯ. Источники полезной энергии в живых существах замечательно однородны. В живых существах для выполнения химического синтеза, сокращения мышечных волокон, активного переноса веществ и для образования полупроницаемых мембран используется энергия солнечного излучения и промежуточного метаболизма, энергия богатых энергией эфирных связей и накопленная энергия восстановленных полимеров. Это использование энергии не имеет видимой связи с электромеханическими преобразователями наших моделей.
в) СИСТЕМА ПОСЛЕДОВАТЕЛЬНОГО УПОРЯДОЧЕНИЯ. У всякого живого существа на некоторой ступени жизненного цикла происходит процесс размножения, а именно митоз. Одноклеточные организмы удваивают свои размеры и затем делятся, причем весь их цикл сводится к митозу. В ходе этого цикла половина элементов переходит от родителя, а другая половина берется из окружающей среды. В многоклеточных организмах митоз приводит к увеличению числа клеток и, в сочетании с дифференциацией тканей, приводит к образованию формы, характерной для данного организма. При этом большинство составных частей потомства берется из среды, и лишь ничтожная их доля поставляется родительской формой.
В серии ВПУ все части "организма" берутся из среды, за исключением ВПУ IV, в котором одна из трех его частей происходит от "родителя". Таким образом, последовательность воспроизведения ВПУ и митоз нельзя непосредственно сравнивать, и общее в них лишь то, что они выполняются в строго определенной последовательности.
При митозе удвоение происходит в момент, трудно прослеживаемый под микроскопом. Видимая часть этого процесса представляет расщепление удвоенного вещества, в частности нуклеиновой кислоты и связанного с ней вещества, в которых у высших организмов заключены планы их структур. Точно так же как порядок пробивок в "хромосомах" моделей ВПУ управляет их сборкой, расположение нуклеотидных компонентов нуклеиновых кислот в хромосомах живых организмов направляет развитие организмов.
Но хромосомы необязательно содержат свой самокопирующий аппарат. Механизм митоза, выполняющий действительное разделение удвоенного набора планов, также должен быть удвоен при воспроизведении. Разные организмы содержат совершенно разные хромосомы, но все они, по-видимому, содержат копирующий аппарат одного и того же рода с незначительными изменениями. Этот экстрахромосомный аппарат представляет, вероятно, биологический аналог всего "организма" ВПУ. Хромосомы, вероятно, просто служат "перфокартами". Они очень сложны у высших организмов, но представляют очень простую часть в конструкции машин, подобных ВПУ I-V, являющихся простым собранием сложных частей.
Сравнение реальной последовательности элементов в биохимических системах с последовательностью элементов в электромеханических системах должно быть очень плодотворным, хотя в настоящее время этот вопрос мало изучен. Связи между действующими элементами ВПУ осуществляют электрические провода; в живых организмах - диффундирующие ионы и белки.
Механические связи имеются в тех и других системах. Биохимическим аналогом реле, представляющим, по существу, усилитель да-нет, является автокаталитическая ферментная система. Отрицательная обратная связь в биохимических системах обычно осуществляется системами на основе простого или конкурентного торможения. Аналог контакта или контактной пары в ВПУ, вероятно, можно найти в биологическом пороговом эффекте, таком, как возбуждение нервов при превышении потенциала покоя. Ответы на проблемы биохимической сборки нужно искать в таких системах, имеющих надлежащее сходство с упомянутыми самовоспроизводящимися системами. Хотя эта аналогия рискована, она наводит на размышления. Если подобные соображения укажут нам, где нужно искать или как описывать явления жизни, проектирование и изготовление наших моделей имело смысл.
ИНФОРМАЦИОННАЯ ЕМКОСТЬ ВПУ И ПРОИСХОЖДЕНИЕ ЖИЗНИ
Наиболее интересным из динамических процессов воспроизведения является перенос информации от планов к организму. Этот перенос планов от хромосомы или перфокарты в организм, являющийся одним из главных нерешенных вопросов биологии, представляет основу истинного понимания жизни. В электромеханических моделях перенос происходит путем воздействия отверстий в перфокартах на работающие части аппарата. Очевидно, хромосомы являются аналогичными устройствами хранения информации. Поскольку хромосомы описывают организм, они должны быть столь же сложны, как и организм.
Сложность чего угодно уместно характеризовать при помощи теории информации. Легко вычислить сложность "хромосом" модели ВПУ в двоичных единицах (битах); она оказывается небольшой. Сложность реальной хромосомы также можно оценить (постольку, поскольку установлена ее структура). Она очень велика. Каждый раз, когда на перфокарте производится двоичный выбор (пробивать отверстие или не пробивать), требуется одна двоичная единица. В "хромосоме" ВПУ I имеется шесть возможных позиций для пробивок. Из них пробиваются только две. Наличие шести позиций для перфорирования приводит к тому, что "хромосома" должна дать модели шесть двоичных единиц информации, необходимых для ее "восприятия" и выполнения. Однако можно заметить, что модель должна быть "настроена" на информацию, даваемую картой, что достигается только благодаря сложности конструкции. Подобная картина наблюдается и применительно к реальным хромосомам. Здесь роль пуансонов выполняют нуклеотиды, выбранные из возможных нуклеотидов (обычно четырех). Такой четверичный выбор эквивалентен двум двоичным, т.е. одному биту информации на нуклеотид. В модели ДНК (дезоксирибонуклеиновой кислоты) Крика-Уотсона наложены ограничения на нуклеиновые кислоты и допущено только около половины выборов по сравнению с совершенно случайной цепью; поэтому количество потенциальной информации в битах только очень приблизительно равно числу нуклеотидов. Это количество, естественно, различно для разных организмов [3]. В бактериальном вирусе с молекулярным весом около 1,2e8 имеется около 4e5 нуклеотидов (хотя вирус, вероятно, не представляет собой организм). В одноклеточном животном, если принять, что вся его ДНК сосредоточена в хромосомоподобном материале, имеется около 5e6 нуклеотидов; в клетке млекопитающего - около 1e10.
Интересно вычислить сложность для всех моделей ВПУ. Информация, связанная с их собиранием, мала - шесть бит для ВПУ I и девять троичных единиц или около 14 бит для ВПУ IV. Но сами тележки весьма сложны. Общая сложность моделей определяется главным образом сложностью тележек, тогда как в общей сложности организмов основную, если не подавляющую часть, составляет сложность хромосом. Общая сложность ВПУ I может дать некоторое указание порядка величины возможной минимальной сложности органической самовоспроизводящейся системы. *** Это вычисление сложности было произведено путем прямого применения принципов теории информации к рассматриваемому механизму [4]. Рассмотрение механических и электрических соединений между элементами ВПУ I позволяет вычислить сложность всего устройства; она равна 116 бит. Эту цифру нужно считать минимальной, поскольку она не включает информацию, необходимую для "стаскивания" частей из окружающей среды. Тем не менее она показывает, что если имеется свободный приток составных элементов, то возможно существование самовоспроизводящегося устройства с информационной сложностью не выше указанной. Это малое количество информации представляется ничтожным перед огромной потенциальной информационной сложностью живых существ, которую можно определить из числа нуклеотидов в "планах" живых клеток. Но эта величина может характеризовать минимум информации, необходимый для осуществления митоза, своего рода sine qua non (необходимое условие) для воспроизведения живого существа.
Аналогичное вычисление было произведено для ВПУ IV, и количество информации, необходимой для организации системы, оказалось равным приблизительно 570 бит. Хотя удвоение планов в настоящее время представляет общее свойство живых организмов, необязательно предполагать, что самые первые из них удваивали свои планы. В самом деле, при столь простых планах, как планы организмов, состоящих из немногих частей, они могли использовать планы, имевшиеся в окружающей среде, так же как ВПУ I использует планы, имеющиеся в одной из его частей. Барричелли [5, 6] выдвинул замечательную гипотезу, что современная жизнь возникла как "симбиоз" между организмом, уже обладавшим способностью к митозу, соответствующим ВПУ I, и некоторой вирусоподобной системой, планы которой заключены в молекулах нуклеиновой кислоты. Первая дала минимальную самовоспроизводящуюся систему со сложностью порядка нескольких сот бит, а последняя, будучи включена в эту систему, обеспечила необходимую гибкость и изменчивость, которая вызвала эволюцию к более высоким формам. В настоящее время эта идея является чистой фантазией, но она заслуживает внимательного рассмотрения некоторых вытекающих из нее положений. Сейчас принято считать, что самовоспроизводящиеся устройства, как электромеханические, так и биохимические, можно характеризовать полученными таким образом значениями информационной сложности. Это, естественно, дает основание для сравнения этих величин для ВПУ I и ВПУ IV с величинами, которые могли бы потребоваться для самопроизвольного возникновения живых форм.
Были произведены оценки [7, 8] допустимой сложности живых организмов, которые показывают, что сложность первых организмов не могла превышать 100-200 бит. Существо более сложное, например равное по сложности ВПУ IV, вряд ли могло бы возникнуть на земном шаре при первом зарождении жизни. Таким образом, результаты рассмотрения моделей, выводы теории информации и приложения термодинамики, по-видимому, дают право сделать заключение о том, что сложность первых живых существ, вероятно, была весьма мала.
Целесообразно дальнейшие исследования проблемы происхождения жизни с информационной точки зрения развернуть по следующим направлениям: 1) оценка количества "составных частей", которые должны быть взяты из среды; 2) отыскание в биохимических механизмах аналогов, соответствующих переключателям, датчикам, запоминающим и другим устройствам, и 3) разработка конкретных моделей самопроизводящих систем с использованием этих биохимических механизмов. Вычисление свободной энергии, необходимой для функционирования моделей этого класса, позволит определить, насколько вероятно самовоспроизвольное развитие таких систем и можно ли принять какую-либо априорную гипотезу о происхождении жизни.
ОПРЕДЕЛЕНИЕ ПОНЯТИЯ ВОСПРОИЗВЕДЕНИЯ
Мы описали пять очевидных моделей процесса воспроизведения, не определив сам процесс. При проектировании моделей было принято следующее неявное определение: воспроизведение есть действие упорядоченной индивидуальной структуры, которая, входя в некоторую систему, выбирает из окружающей среды случайно расположенные части и строит из них и из своих собственных частей функционально-эквивалентные структуры. Серия моделей ВПУ, очевидно, соответствует этому определению и, по существу, подсказывает его. Это определение исключает процессы, часто называемые процессами воспроизведения, но по интуитивному восприятию не являющиеся таковыми. Например, действия печатной машины или штамповальной машины не подходят под это определение, если только они не производят новые печатные или штамповальные машины.
При проектировании модели воспроизведения необходимо избегать двух опасностей: во-первых, опасности создать тривиально простую машину и, во-вторых, опасности создать нереализуемо сложную систему.
Для ВПУ были выбраны два рода деталей. Это, очевидно, наименьшее число, которое может быть введено в окружающую среду случайно. Число родов деталей можно свести к одному, но тогда не будет выполнено требование случайности. ВПУ I при помощи любопытного процесса можно свести к одноэлементной системе. При этом тележки А ставят на место, по одной на ветке, и устанавливают так, что их "хромосомы" находятся во второй позиции, т.е. их инструкции позволяют входить на ветку только тележке В. Если одна тележка В впускается на первую ветку, последующие ветки заполняются одна за другой тележками В. Хотя это прекрасный пример автоматического последовательного устройства, это уже не будет представлять воспроизведения.
Такой процесс напоминает кристаллизацию, которую также часто приводят как пример воспроизведения. Несомненно, при кристаллизации происходит усложнение структуры, однако в нем нет отчетливой индивидуализации. Ее нет даже в такой двухкомпонентной кристаллизующейся системе, как хлористый натрий. О таком кристалле никак нельзя сказать, что данный ион натрия или хлора произошел от какого-нибудь другого конкретного иона или данной группы ионов.
Часто в качестве аналога процесса воспроизведения приводят другую химическую параллель - автокатализ. Автокатализ также отличается от воспроизведения отсутствием индивидуальности. Для объяснения возникновения жизни, которое часто связывают с процессом кристаллизации или автокатализа, требуется большее, а именно объяснить происхождение индивидуальности, которая выражается в сходстве между родителями и потомками, наблюдаемом во всей живой природе. Некоторые виды кристаллизации и автокатализа были, без сомнения, предвестниками жизни, однако они не удовлетворяют нашему определению воспроизведения.
Определение неполно и в том отношении, что оно не указывает характер окружающей среды, допускающей воспроизведение. По-видимому, она может быть произвольной. Две абстрактные модели воспроизведения были упомянуты фон Нейманом [1, 2], который первый обосновал возможность создания подобных автоматов. В этих моделях в качестве первого типа окружающей среды была указана внутренняя структура перфорационной машины Тьюринга - мало вероятный случай среды; второй тип среды - своеобразная группа смежных прямоугольных клеток, причем изменения в одной клетке могут воздействовать на конечное число состояний в соседней клетке. Оба эти типа среды весьма произвольны, как и в случае моделей ВПУ. Единственное, что делает модели ВПУ шагом вперед по сравнению с системами фон Неймана - то, что они спроектированы в деталях, и по крайней мере одна из них работает.
Неприменимость таких произвольных типов среды заключается в их искусственности. Биологи ищут систему, действующую в среде, не требующей вмешательства человека, т.е. модель самопроизвольного возникновения самовоспроизводящегося вида. Жизнь, конечно, появилась не на рельсовом пути макета железной дороги! Но наши тележки следует принимать как модели, независимо от типа среды. По-видимому, их можно расценивать по изяществу, которое будет больше в моделях с более простым окружением, с меньшей аппаратурой и более простыми частями.
Мы осмеливаемся теперь высказать мысль, что этот процесс самоупорядочения можно ввести в электронную систему. Хотя в ней не будет наглядности движущихся частей и игрушечных поездов, устройство из триггерных схем, магнитных элементов или реле может быть сделано столь же близким к нашему определению, как любые из упомянутых моделей. Если построить системы электрических цепей таким образом, что упорядочение одной известным образом заставляет другие упорядочиваться таким же образом, то мы получим электронную модель воспроизведения.
АНАЛОГИ БИОЛОГИЧЕСКИХ СИСТЕМ И ИХ ФУНКЦИЙ
Ценность моделей этого рода состоит в том, что они привлекают наше внимание к абстрактным функциям, присущим моделируемым ими процессам. Модель, как и живое существо, имеет некоторый необходимый минимальный набор функций. Мы уже указали на абсолютную необходимость: а) источника составных частей, двигателя, планов и некоторого механизма опознавания, б) источника энергии и в) системы передачи и обработки информации или последовательного упорядочения. Их можно считать минимальными функциями, самовоспроизводящейся системы. Рассмотрим аналоги каждой из них.
а) СОСТАВНЫЕ ЧАСТИ. В состав живых существ входят вода, соли, аминокислоты, жирные кислоты и витамины для животных; вода, углекислота и соли для растений. Из этих простых частей организмы создают свои копии.
Из сравнения с моделями ВПУ видно, что живые существа представляют собой сложные собрания простых частей, а модели - простые собрания сложных частей. В этом смысле модели не представляют собой подлинные копии живых существ. Представляется удивительным, сколь малым числом составных частей может обойтись простое живое существо, обладая при этом способностью функционировать и воспроизводить самого себя. Это, конечно, зависит от того, что называть "частью". Если исходить из частей, уже имеющихся в готовом виде в окружающей среде, то моделям нужны две или три такие части, а живым существам - миллионы частей. Естественная, реальная, т.е. неискусственная среда, вряд ли содержит значительное число столь сложных частей, как описанные тележки A и B. Вероятность появления таких частей зависит от их сложности, от того, сколь часто их составные части встречаются в окружающей среде, и от свободной энергии устройства. Мы обошли это затруднение, изготовив части заранее. В природе устройства, равные по сложности тележкам A и B, могут не появиться в течение значительного времени. Поэтому вопрос о том, были ли первые живые существа собраниями небольшого числа сложных частей или большого числа простых частей, остается совершенно нерешенным. Всякая попытка сделать выводы по этому пункту может дать лишь весьма убогие результаты.
б) ЭНЕРГИЯ. Источники полезной энергии в живых существах замечательно однородны. В живых существах для выполнения химического синтеза, сокращения мышечных волокон, активного переноса веществ и для образования полупроницаемых мембран используется энергия солнечного излучения и промежуточного метаболизма, энергия богатых энергией эфирных связей и накопленная энергия восстановленных полимеров. Это использование энергии не имеет видимой связи с электромеханическими преобразователями наших моделей.
в) СИСТЕМА ПОСЛЕДОВАТЕЛЬНОГО УПОРЯДОЧЕНИЯ. У всякого живого существа на некоторой ступени жизненного цикла происходит процесс размножения, а именно митоз. Одноклеточные организмы удваивают свои размеры и затем делятся, причем весь их цикл сводится к митозу. В ходе этого цикла половина элементов переходит от родителя, а другая половина берется из окружающей среды. В многоклеточных организмах митоз приводит к увеличению числа клеток и, в сочетании с дифференциацией тканей, приводит к образованию формы, характерной для данного организма. При этом большинство составных частей потомства берется из среды, и лишь ничтожная их доля поставляется родительской формой.
В серии ВПУ все части "организма" берутся из среды, за исключением ВПУ IV, в котором одна из трех его частей происходит от "родителя". Таким образом, последовательность воспроизведения ВПУ и митоз нельзя непосредственно сравнивать, и общее в них лишь то, что они выполняются в строго определенной последовательности.
При митозе удвоение происходит в момент, трудно прослеживаемый под микроскопом. Видимая часть этого процесса представляет расщепление удвоенного вещества, в частности нуклеиновой кислоты и связанного с ней вещества, в которых у высших организмов заключены планы их структур. Точно так же как порядок пробивок в "хромосомах" моделей ВПУ управляет их сборкой, расположение нуклеотидных компонентов нуклеиновых кислот в хромосомах живых организмов направляет развитие организмов.
Но хромосомы необязательно содержат свой самокопирующий аппарат. Механизм митоза, выполняющий действительное разделение удвоенного набора планов, также должен быть удвоен при воспроизведении. Разные организмы содержат совершенно разные хромосомы, но все они, по-видимому, содержат копирующий аппарат одного и того же рода с незначительными изменениями. Этот экстрахромосомный аппарат представляет, вероятно, биологический аналог всего "организма" ВПУ. Хромосомы, вероятно, просто служат "перфокартами". Они очень сложны у высших организмов, но представляют очень простую часть в конструкции машин, подобных ВПУ I-V, являющихся простым собранием сложных частей.
Сравнение реальной последовательности элементов в биохимических системах с последовательностью элементов в электромеханических системах должно быть очень плодотворным, хотя в настоящее время этот вопрос мало изучен. Связи между действующими элементами ВПУ осуществляют электрические провода; в живых организмах - диффундирующие ионы и белки.
Механические связи имеются в тех и других системах. Биохимическим аналогом реле, представляющим, по существу, усилитель да-нет, является автокаталитическая ферментная система. Отрицательная обратная связь в биохимических системах обычно осуществляется системами на основе простого или конкурентного торможения. Аналог контакта или контактной пары в ВПУ, вероятно, можно найти в биологическом пороговом эффекте, таком, как возбуждение нервов при превышении потенциала покоя. Ответы на проблемы биохимической сборки нужно искать в таких системах, имеющих надлежащее сходство с упомянутыми самовоспроизводящимися системами. Хотя эта аналогия рискована, она наводит на размышления. Если подобные соображения укажут нам, где нужно искать или как описывать явления жизни, проектирование и изготовление наших моделей имело смысл.
ИНФОРМАЦИОННАЯ ЕМКОСТЬ ВПУ И ПРОИСХОЖДЕНИЕ ЖИЗНИ
Наиболее интересным из динамических процессов воспроизведения является перенос информации от планов к организму. Этот перенос планов от хромосомы или перфокарты в организм, являющийся одним из главных нерешенных вопросов биологии, представляет основу истинного понимания жизни. В электромеханических моделях перенос происходит путем воздействия отверстий в перфокартах на работающие части аппарата. Очевидно, хромосомы являются аналогичными устройствами хранения информации. Поскольку хромосомы описывают организм, они должны быть столь же сложны, как и организм.
Сложность чего угодно уместно характеризовать при помощи теории информации. Легко вычислить сложность "хромосом" модели ВПУ в двоичных единицах (битах); она оказывается небольшой. Сложность реальной хромосомы также можно оценить (постольку, поскольку установлена ее структура). Она очень велика. Каждый раз, когда на перфокарте производится двоичный выбор (пробивать отверстие или не пробивать), требуется одна двоичная единица. В "хромосоме" ВПУ I имеется шесть возможных позиций для пробивок. Из них пробиваются только две. Наличие шести позиций для перфорирования приводит к тому, что "хромосома" должна дать модели шесть двоичных единиц информации, необходимых для ее "восприятия" и выполнения. Однако можно заметить, что модель должна быть "настроена" на информацию, даваемую картой, что достигается только благодаря сложности конструкции. Подобная картина наблюдается и применительно к реальным хромосомам. Здесь роль пуансонов выполняют нуклеотиды, выбранные из возможных нуклеотидов (обычно четырех). Такой четверичный выбор эквивалентен двум двоичным, т.е. одному биту информации на нуклеотид. В модели ДНК (дезоксирибонуклеиновой кислоты) Крика-Уотсона наложены ограничения на нуклеиновые кислоты и допущено только около половины выборов по сравнению с совершенно случайной цепью; поэтому количество потенциальной информации в битах только очень приблизительно равно числу нуклеотидов. Это количество, естественно, различно для разных организмов [3]. В бактериальном вирусе с молекулярным весом около 1,2e8 имеется около 4e5 нуклеотидов (хотя вирус, вероятно, не представляет собой организм). В одноклеточном животном, если принять, что вся его ДНК сосредоточена в хромосомоподобном материале, имеется около 5e6 нуклеотидов; в клетке млекопитающего - около 1e10.
Интересно вычислить сложность для всех моделей ВПУ. Информация, связанная с их собиранием, мала - шесть бит для ВПУ I и девять троичных единиц или около 14 бит для ВПУ IV. Но сами тележки весьма сложны. Общая сложность моделей определяется главным образом сложностью тележек, тогда как в общей сложности организмов основную, если не подавляющую часть, составляет сложность хромосом. Общая сложность ВПУ I может дать некоторое указание порядка величины возможной минимальной сложности органической самовоспроизводящейся системы. *** Это вычисление сложности было произведено путем прямого применения принципов теории информации к рассматриваемому механизму [4]. Рассмотрение механических и электрических соединений между элементами ВПУ I позволяет вычислить сложность всего устройства; она равна 116 бит. Эту цифру нужно считать минимальной, поскольку она не включает информацию, необходимую для "стаскивания" частей из окружающей среды. Тем не менее она показывает, что если имеется свободный приток составных элементов, то возможно существование самовоспроизводящегося устройства с информационной сложностью не выше указанной. Это малое количество информации представляется ничтожным перед огромной потенциальной информационной сложностью живых существ, которую можно определить из числа нуклеотидов в "планах" живых клеток. Но эта величина может характеризовать минимум информации, необходимый для осуществления митоза, своего рода sine qua non (необходимое условие) для воспроизведения живого существа.
Аналогичное вычисление было произведено для ВПУ IV, и количество информации, необходимой для организации системы, оказалось равным приблизительно 570 бит. Хотя удвоение планов в настоящее время представляет общее свойство живых организмов, необязательно предполагать, что самые первые из них удваивали свои планы. В самом деле, при столь простых планах, как планы организмов, состоящих из немногих частей, они могли использовать планы, имевшиеся в окружающей среде, так же как ВПУ I использует планы, имеющиеся в одной из его частей. Барричелли [5, 6] выдвинул замечательную гипотезу, что современная жизнь возникла как "симбиоз" между организмом, уже обладавшим способностью к митозу, соответствующим ВПУ I, и некоторой вирусоподобной системой, планы которой заключены в молекулах нуклеиновой кислоты. Первая дала минимальную самовоспроизводящуюся систему со сложностью порядка нескольких сот бит, а последняя, будучи включена в эту систему, обеспечила необходимую гибкость и изменчивость, которая вызвала эволюцию к более высоким формам. В настоящее время эта идея является чистой фантазией, но она заслуживает внимательного рассмотрения некоторых вытекающих из нее положений. Сейчас принято считать, что самовоспроизводящиеся устройства, как электромеханические, так и биохимические, можно характеризовать полученными таким образом значениями информационной сложности. Это, естественно, дает основание для сравнения этих величин для ВПУ I и ВПУ IV с величинами, которые могли бы потребоваться для самопроизвольного возникновения живых форм.
Были произведены оценки [7, 8] допустимой сложности живых организмов, которые показывают, что сложность первых организмов не могла превышать 100-200 бит. Существо более сложное, например равное по сложности ВПУ IV, вряд ли могло бы возникнуть на земном шаре при первом зарождении жизни. Таким образом, результаты рассмотрения моделей, выводы теории информации и приложения термодинамики, по-видимому, дают право сделать заключение о том, что сложность первых живых существ, вероятно, была весьма мала.
Целесообразно дальнейшие исследования проблемы происхождения жизни с информационной точки зрения развернуть по следующим направлениям: 1) оценка количества "составных частей", которые должны быть взяты из среды; 2) отыскание в биохимических механизмах аналогов, соответствующих переключателям, датчикам, запоминающим и другим устройствам, и 3) разработка конкретных моделей самопроизводящих систем с использованием этих биохимических механизмов. Вычисление свободной энергии, необходимой для функционирования моделей этого класса, позволит определить, насколько вероятно самовоспроизвольное развитие таких систем и можно ли принять какую-либо априорную гипотезу о происхождении жизни.
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Чт Май 20, 2021 1:28 am
ПРИЛОЖЕНИЕ А. УСТРОЙСТВО И КОНСТРУКЦИЯ МОДЕЛИ ВПУ I
На фото I-VII изображена работающая модель ВПУ I. Фотографии I-III воспроизводят общий вид модели, а - IV-VII - подробности и взаимодействие тележек.

Фото I. Общая схема ВПУ I, вид сверху.
На фото I показан общий вид установки без тележек. Видны дорожка "среды" и четыре ветки с тупиками, а также три пары кнопок, управляющих стрелками этих трех веток, и два миниатюрных переключателя, управляющих описываемой ниже "системой задержки" тележек.

Фото II. ВПУ I в начале цикла. Собран первый "организм".
На фото II изображен прибор в перспективе. Тележки со словами "голова" и "хвост" (head, tail) расположены случайно, в порядке, полученном в результате бросания монеты, таким образом: ГХГГХХХГГХ, и первые две тележки направлены на первую ветку. Сборка первого "организма" происходила под руководством оператора, управляющего двумя кнопками на главном блоке управления, расположенном направо. Кнопки выключают и включают стрелку первой ветки, так называемую "стрелку Провидения". "Источник питания" размещен наверху слева.

Фото III. ВПУ I в конце цикла. Собраны все четыре "организма"; для оставшихся частей нет места.
На фото III показано прекращение "вида", когда на всех четырех ветках находятся бездействующие "организмы". Дальнейшее "воспроизведение" невозможно, так как больше нет свободных веток, хотя в окружающей "среде" - на дорожке - имеется еще одна тележка А ("голова") и В ("хвост").

Фото IV. Собранный "организм".
Прибор работает на постоянном токе при напряжении 10-12 вольт, которое подводится к рельсам. Он установлен на фанерной кленовой панели площадью в 215*60см с трубчатыми стальными ножками. На фото IV изображены детали одной пары тележек A и B, т.е. единичного "организма". Тележки состоят из литой рамы, на которой спереди установлен обычный двигатель, а сзади два изолированных колеса на бакелитовом бруске. На тележках видны реле, контакты, рычаги и довольно сложная система с храповичком на тележке A. Тележки, как видно на рисунке, останавливаются тупиком на ветке, а электрическое соединение между ними создают четыре буферных контакта.

Фото V. Тележки A и B с буферными контактами.
На фото V можно видеть яснее конструкцию тележек и четырех контактов.

Фото VI. Приближение тележки B к "организму" обнаружено "глазом B" на тележке B собранного "организма".

Фото VII. Прохождение тележки B на ветку обнаружено "глазом A или B" на тележке B собранного "организма".
На фото VI показано приближение тележки B к хвостовой части "организма", собранного на ветке, а на фото VII показано взаимодействие между тележкой B, входящей на ветку, и тележкой B уже собранного "организма" на предшествующей ветке.
Два реле на тележках В, которые особенно ясно видны на фото IV, возбуждают стрелки, управляющие следующей веткой. Удлиненный рычаг, установленный на подвижной крышке реле, имеет на конце контакт, в который поступает ток от реле. При замыкании реле контакт опускается до полотна дороги. Под рычагом расположены кнопки, через которые ток проходит к стрелке. При замыкании заднего реле стрелка открывает следующую ветку, а при замыкании переднего реле стрелка закрывает эту ветку.

Рис.5. Контактные "глаза" тележки B и рычаги тележек A и B.
Через реле должен протекать ток надлежащего направления. Направление тока определяется тем, что "организм" "знает", какая тележка проходит мимо него к следующей ветке. Это значит, что тележка должна выглядеть по-разному для некоего "органа" на тележке Б, на которой установлены не только "мышцы" - реле, но и контактные "глаза". "Глаза" представляют пару различно расположенных контактов; один из них "чувствителен" к прохождению тележкам A, а другой - к тележкам B. Чувствительность обеспечивается тем, что на тележках A и B установлены опознавательные рычаги, которые доходят до ветки и замыкают соответствующий контакт на тележке B "организма", "сообщая" тем самым, принадлежат они к типу A или B. Это видно на фото VI и особенно ясно на рис.5. Рычаг на тележке A расположен на 6мм выше, чем рычаг на тележке B, и поэтому не касается нижней пары контактов; рычаг на тележке B не доходит до верхней пары контактов, потому что он расположен на сантиметр ниже рычага тележки A. Поэтому при прохождении тележки A замыкается верхняя (дальняя) пара контактов. При соответствующей настройке электрической схемы "организма" замыкание пары контактов вызовет прохождение тока через хвостовое реле к тележке B, и стрелка откроет следующую ветку. То же самое может произойти при прохождении тележки B. При этом замыкается нижняя (ближняя) пара контактов. Эти пары контактов мы будем называть соответственно "глаза A" и "глаза B". На рычагах написано "голова" и "хвост".
Такого же рода операции должны выполняться при управлении закрыванием стрелки следующей ветки. Закрывание должно происходить после того, как тележка прошла стрелку. Его можно было бы сделать автоматическим, но в данной модели это не было сделано.
На правой стороне тележки B установлена третья пара контактов "глаз А или В". Она лучше всего видна на фото VII. "Глаза A или В" представляют собой пару изогнутых металлических полосок посередине боковой стороны тележек. Эти "глаза" реагируют на короткий выступ с левой стороны тележек A или B, который можно видеть на фото VII. Как только тележки A или B пройдут за стрелку, эта пара контактов замыкается. При замыкании контактов ток проходит к переднему рычагу на тележке B, который "закрывает" стрелку на следующую ветку.
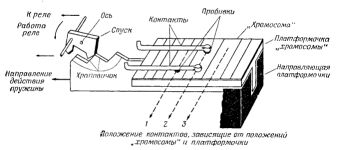
Рис.6. Детали механизма передвижения "хромосомы".
Рассмотрим вопрос об установлении последовательности действий, которые должны выполнять "глаза". Сначала они должны быть чувствительны к приближению только тележки A; после того как тележка A прошла на ветку, они должны стать чувствительными только к тележке В; а после того как и тележка B прошла на ветку, они должны потерять всякую чувствительность к тележкам. Этой последовательностью управляет храповичок на тележке A, или "мозг". На рис.6 схематически показаны детали механизма. Он состоит из реле, храповичка и "хромосомы" - перфорированной карты с кодированными командами, двух контактов и пружинной платформочки, установленной на направляющей, скользящей по канавке в раме тележки. При возбуждении реле перемещает храповичок вверх и вниз, так что платформочка "хромосомы" продвигается по направляющей на ход одного зубца храповичка (6мм). Ввиду того что "хромосома" является изолятором, отделяющим контакты от проводящей платформочки и направляющей, ее движение вызывает периодический разрыв цепи между контактами и направляющей.

Рис.7. Электрическая схема тележек ВПУ I.
В начале "жизненного цикла" один контакт (нижний контакт на рис.6 или контакт Cr1 на рис.7) соединяется с направляющей через первое отверстие хромосомы. Этот контакт соединен с контактной парой "глаза" A на тележке B через один из буферных контактов между двумя тележками. В этом положении он заземляет один из контактов, сообщая "организму" "чувствительность" к прохождению тележек A на дорожке окружающей среды.
Когда тележка A входит на ветку, это вызывает возбуждение реле, управляющего "хромосомой", и заставляет ее передвинуться на один зубец вместе с платформочкой. Теперь она находится в положении, изображенном на рис.6, а контакты находятся во втором положении. При этом другой контакт (верхний контакт на рис.6 или контакт Cr2 на рис.7) соединен с направляющей через второе отверстие в "хромосоме", а первый контакт изолирован от направляющей. Другая контактная пара (контакты "глаза" B на тележке B) становится чувствительной к прохождению тележек B, и один контакт этой пары заземляется. Далее тележка B направляется на ветку, а "хромосома" и ее платформочка опять передвигаются на один зубец под действием своего реле. Теперь контакты находятся в положении 3, как показано на рис.6, и оба контакта изолированы от земли неперфорированным участком "хромосомы". Вследствие этого дальнейшее продвижение тележек на ветку не происходит, так как оба контактные "глаза" на тележке B изолированы. Поскольку на ветке образовался полный "организм", ему не нужно производить дальнейших переключений.
Интересно рассмотреть, что получится при изменении положения пробивок в "хромосоме". При очень небольших изменениях "организм" может получать от "хромосомы" любой набор из трех команд. Одна "хромосома" была перфорирована так, что она подавала стрелкам команды в обратном порядке, т.е. "организм" собирался на ветке в обратном порядке. Естественно, что такая "мутация" совершенно нежизненна, так как при этом не произойдет "воспроизведения". Можно считать, что тележка, снабженная такими обратными командами, имеет мутировавший "ген", выраженный в "фенотипе" следующего "поколения". Такие неправильные приспособления и были установлены на тележке A, которую можно заметить в группе, изображенной на фото III, на четвертой ветке (крайней справа) и на которой надпись "head" ("голова") перевернута, чтобы показать, что "мозг" этой тележки инвертирован.
На рис.7 показана полная схема всех реле, контактов, "хромосом" и стрелок для веток и тележек ВПУ I. Обозначения элементов схемы следующие. На тележке A:
1. "Хромосома" и направляющая Cr с отверстиями, обозначенными A и B. Направляющая на рисунке заземлена.
2. Контакты Cr1 и Cr2, проводящие команды от "хромосомы" к тележке B.
3. Реле "хромосомы" R; механическая связь этого реле с Cr показана пунктирной линией со стрелкой. Реле попеременно заземляет Cr1 и Cr2 и затем окончательно отключает их от земли.
4. Четыре буферных контакта C1-C4, соединяющие тележки A и B.
5. Рычаг A (тележки A) и выступ F.
На тележке B:
1. Пара реле, управляющих стрелками веток, R1 и R2. Реле R1, будучи возбуждено, подает ток на переключатель открывания ветки S1, и стрелка открывает ветку. При возбуждении реле R2 ток поступает на переключатель замыкания ветки S2, и ветка закрывается.
2. Пара контактов "глаз" ЕА, чувствительных к рычагу A (тележки A); пара контактов "глаз" ЕB, чувствительных к рычагу A (тележки B); обе пары "глаз" реагируют на тележки, проходящие по дорожке среды.
3. Пара контактов ЕАВ или "глаз А или В", реагирующий на выступ на тележке, когда она проходит на следующую ветку.
4. Четыре буферных контакта C1-C4.
5. Рычаг А (тележки В) и выступ F.
Кроме того, на обеих тележках стоят двигатели и источники питания, получающие энергию от рельс. Двигатель тележки А выключается, когда тележка доходит до конца ветки, вследствие размыкания цепи от положительного полюса блока питания на полотне ветки. Но двигатель тележки В продолжает работать и движение вызывает давление в контактах, обеспечивающее надежное соединение между тележками.
Последовательность операций, приводящая к сборке организмов, следующая:
1. Оператор, действуя органами управления "стрелки Провидения" на главном пульте управления, собирает "организм" на первой ветке.
2. Буферный контакт C1 заземляет контакты EA через платформочку Cr1.
3. Прохождение тележки A по дорожке среды вызывает замыкание контактов EA под действием рычага A (тележки A).
4. Вследствие этого возбуждается реле открывания ветки R1, и переключатель S1 открывает ветку.
5. Тележка входит на вторую ветку.
6. Когда тележка А проходит стрелку второй ветки, действие выступа F на входящей тележке вызывает замыкание контактов "глаза" EAB.
7. Замыкание контактоз EAB вызывает возбуждение реле закрывания ветки R2, которое закрывает ветку посредством переключателя S2.
8. Одновременно с (7) замыкание контактов EAB вызывает возбуждение реле "хромосомы" R через платформочку C3. "Хромосома" Cr благодаря этому передвигается вперед во второе положение.
9. Вследствие этого один полюс контактов "глаза" EB заземляется через C2 и Cr2.
10. Прохождение тележки B по дорожке среды вызывает замыкание контактов EB под действием рычага A (тележки B).
11. Вследствие этого возбуждается реле открывания ветки R1, заставляющее переключатель S1 открыть ветку.
12. Тележка B входит на вторую ветку.
13. Когда тележка B проходит через стрелку второй ветки, выступом F на тележке B замыкаются контакты "глаза" EAB.
14. Замыкание контактов EAB вызывает возбуждение реле ветки R2, закрывающее ветку посредством переключателя S2.
15. Одновременно с (14) замыкание контактов EAB вызывает возбуждение реле "хромосомы" R через C3. Вследствие этого "хромосома" Cr передвигается в третье положение.
16. При третьем положении "хромосомы" Cr "глаза" EA и EB, а также Cr и Cr2, отключаются от земли, и вследствие этого "организм" на первой ветке больше не "воспринимает" и не "передает" информации.
17. "Организм" на второй ветке повторяет этапы (2-16) по отношению к третьей ветке; процесс продолжается до тех пор, пока все ветки не будут заполнены или не будут исчерпаны все запасы частей или не произойдет какая-нибудь ошибка.
Случайное столкновение тележек A и B на дорожке среды не имеет значения, так как тележки могут действовать, только находясь на ветке, и положение "хромосомы" может измениться только тогда, когда она пропускает тележку на следующую ветку.
Оказалось абсолютно необходимым принятие одной предосторожности, а именно введение "системы задержки" тележек.
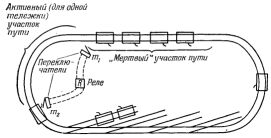
Рис.8. Схема системы задержки.
Если тележки будут свободно двигаться вместе по дорожке среды, они могут подходить друг к другу вплотную. В таком случае, когда" первая тележка будет направлена на ветку, вторая тележка вследствие конечной длины стрелочного механизма обязательно проскочит за ней на ветку, прежде чем стрелка будет закрыта. Это приведет к окончанию цикла. Поэтому было придумано устройство, обеспечивающее, чтобы две тележки не подходили слишком близко одна к другой, когда первая тележка подходит к стрелке. Схема этого устройства изображена на рис.8, и некоторые его элементы можно видеть на фото I-III. Оно представляет участок полотна, которой обесточивается, как только тележка сходит с него и входит на находящийся перед ним активный участок. Только достигнув конца активного участка, расположенного перед стрелками, тележка снова подает ток на "мертвый" участок. Следующая тележка, пройдя "мертвый" участок, в свою очередь обесточит его для следующих тележек. Действие "системы задержки" выражается в том, что тележки выходят с участка поодиночке. Передний конец участка расположен непосредственно перед закругленным активным участком, находящимся перед разветвлениями. Таким образом, когда тележка достигает разветвления, следующая тележка подходит за ней, но не вплотную, так как скорости тележек примерно одинаковы. Устройство служило не только для предупреждения неполадок при сборке, но и для того, чтобы тележки не толкали друг друга на закругленном участке и не сходили с рельс. Действие "системы задержки" можно рассмотреть на фото II, на котором видно, что шесть тележек задержаны на "мертвом" участке пути, одна тележка находится на активном участке и одна готова войти на вторую ветку. Две тележки вошли на первую ветку. Ясно видно, что тележки на закругленном участке и в разветвлениях отделены друг от друга, так как накопление тележек происходит на прямом "мертвом" участке.
На фото I-VII изображена работающая модель ВПУ I. Фотографии I-III воспроизводят общий вид модели, а - IV-VII - подробности и взаимодействие тележек.

Фото I. Общая схема ВПУ I, вид сверху.
На фото I показан общий вид установки без тележек. Видны дорожка "среды" и четыре ветки с тупиками, а также три пары кнопок, управляющих стрелками этих трех веток, и два миниатюрных переключателя, управляющих описываемой ниже "системой задержки" тележек.

Фото II. ВПУ I в начале цикла. Собран первый "организм".
На фото II изображен прибор в перспективе. Тележки со словами "голова" и "хвост" (head, tail) расположены случайно, в порядке, полученном в результате бросания монеты, таким образом: ГХГГХХХГГХ, и первые две тележки направлены на первую ветку. Сборка первого "организма" происходила под руководством оператора, управляющего двумя кнопками на главном блоке управления, расположенном направо. Кнопки выключают и включают стрелку первой ветки, так называемую "стрелку Провидения". "Источник питания" размещен наверху слева.

Фото III. ВПУ I в конце цикла. Собраны все четыре "организма"; для оставшихся частей нет места.
На фото III показано прекращение "вида", когда на всех четырех ветках находятся бездействующие "организмы". Дальнейшее "воспроизведение" невозможно, так как больше нет свободных веток, хотя в окружающей "среде" - на дорожке - имеется еще одна тележка А ("голова") и В ("хвост").

Фото IV. Собранный "организм".
Прибор работает на постоянном токе при напряжении 10-12 вольт, которое подводится к рельсам. Он установлен на фанерной кленовой панели площадью в 215*60см с трубчатыми стальными ножками. На фото IV изображены детали одной пары тележек A и B, т.е. единичного "организма". Тележки состоят из литой рамы, на которой спереди установлен обычный двигатель, а сзади два изолированных колеса на бакелитовом бруске. На тележках видны реле, контакты, рычаги и довольно сложная система с храповичком на тележке A. Тележки, как видно на рисунке, останавливаются тупиком на ветке, а электрическое соединение между ними создают четыре буферных контакта.

Фото V. Тележки A и B с буферными контактами.
На фото V можно видеть яснее конструкцию тележек и четырех контактов.

Фото VI. Приближение тележки B к "организму" обнаружено "глазом B" на тележке B собранного "организма".

Фото VII. Прохождение тележки B на ветку обнаружено "глазом A или B" на тележке B собранного "организма".
На фото VI показано приближение тележки B к хвостовой части "организма", собранного на ветке, а на фото VII показано взаимодействие между тележкой B, входящей на ветку, и тележкой B уже собранного "организма" на предшествующей ветке.
Два реле на тележках В, которые особенно ясно видны на фото IV, возбуждают стрелки, управляющие следующей веткой. Удлиненный рычаг, установленный на подвижной крышке реле, имеет на конце контакт, в который поступает ток от реле. При замыкании реле контакт опускается до полотна дороги. Под рычагом расположены кнопки, через которые ток проходит к стрелке. При замыкании заднего реле стрелка открывает следующую ветку, а при замыкании переднего реле стрелка закрывает эту ветку.

Рис.5. Контактные "глаза" тележки B и рычаги тележек A и B.
Через реле должен протекать ток надлежащего направления. Направление тока определяется тем, что "организм" "знает", какая тележка проходит мимо него к следующей ветке. Это значит, что тележка должна выглядеть по-разному для некоего "органа" на тележке Б, на которой установлены не только "мышцы" - реле, но и контактные "глаза". "Глаза" представляют пару различно расположенных контактов; один из них "чувствителен" к прохождению тележкам A, а другой - к тележкам B. Чувствительность обеспечивается тем, что на тележках A и B установлены опознавательные рычаги, которые доходят до ветки и замыкают соответствующий контакт на тележке B "организма", "сообщая" тем самым, принадлежат они к типу A или B. Это видно на фото VI и особенно ясно на рис.5. Рычаг на тележке A расположен на 6мм выше, чем рычаг на тележке B, и поэтому не касается нижней пары контактов; рычаг на тележке B не доходит до верхней пары контактов, потому что он расположен на сантиметр ниже рычага тележки A. Поэтому при прохождении тележки A замыкается верхняя (дальняя) пара контактов. При соответствующей настройке электрической схемы "организма" замыкание пары контактов вызовет прохождение тока через хвостовое реле к тележке B, и стрелка откроет следующую ветку. То же самое может произойти при прохождении тележки B. При этом замыкается нижняя (ближняя) пара контактов. Эти пары контактов мы будем называть соответственно "глаза A" и "глаза B". На рычагах написано "голова" и "хвост".
Такого же рода операции должны выполняться при управлении закрыванием стрелки следующей ветки. Закрывание должно происходить после того, как тележка прошла стрелку. Его можно было бы сделать автоматическим, но в данной модели это не было сделано.
На правой стороне тележки B установлена третья пара контактов "глаз А или В". Она лучше всего видна на фото VII. "Глаза A или В" представляют собой пару изогнутых металлических полосок посередине боковой стороны тележек. Эти "глаза" реагируют на короткий выступ с левой стороны тележек A или B, который можно видеть на фото VII. Как только тележки A или B пройдут за стрелку, эта пара контактов замыкается. При замыкании контактов ток проходит к переднему рычагу на тележке B, который "закрывает" стрелку на следующую ветку.

Рис.6. Детали механизма передвижения "хромосомы".
Рассмотрим вопрос об установлении последовательности действий, которые должны выполнять "глаза". Сначала они должны быть чувствительны к приближению только тележки A; после того как тележка A прошла на ветку, они должны стать чувствительными только к тележке В; а после того как и тележка B прошла на ветку, они должны потерять всякую чувствительность к тележкам. Этой последовательностью управляет храповичок на тележке A, или "мозг". На рис.6 схематически показаны детали механизма. Он состоит из реле, храповичка и "хромосомы" - перфорированной карты с кодированными командами, двух контактов и пружинной платформочки, установленной на направляющей, скользящей по канавке в раме тележки. При возбуждении реле перемещает храповичок вверх и вниз, так что платформочка "хромосомы" продвигается по направляющей на ход одного зубца храповичка (6мм). Ввиду того что "хромосома" является изолятором, отделяющим контакты от проводящей платформочки и направляющей, ее движение вызывает периодический разрыв цепи между контактами и направляющей.

Рис.7. Электрическая схема тележек ВПУ I.
В начале "жизненного цикла" один контакт (нижний контакт на рис.6 или контакт Cr1 на рис.7) соединяется с направляющей через первое отверстие хромосомы. Этот контакт соединен с контактной парой "глаза" A на тележке B через один из буферных контактов между двумя тележками. В этом положении он заземляет один из контактов, сообщая "организму" "чувствительность" к прохождению тележек A на дорожке окружающей среды.
Когда тележка A входит на ветку, это вызывает возбуждение реле, управляющего "хромосомой", и заставляет ее передвинуться на один зубец вместе с платформочкой. Теперь она находится в положении, изображенном на рис.6, а контакты находятся во втором положении. При этом другой контакт (верхний контакт на рис.6 или контакт Cr2 на рис.7) соединен с направляющей через второе отверстие в "хромосоме", а первый контакт изолирован от направляющей. Другая контактная пара (контакты "глаза" B на тележке B) становится чувствительной к прохождению тележек B, и один контакт этой пары заземляется. Далее тележка B направляется на ветку, а "хромосома" и ее платформочка опять передвигаются на один зубец под действием своего реле. Теперь контакты находятся в положении 3, как показано на рис.6, и оба контакта изолированы от земли неперфорированным участком "хромосомы". Вследствие этого дальнейшее продвижение тележек на ветку не происходит, так как оба контактные "глаза" на тележке B изолированы. Поскольку на ветке образовался полный "организм", ему не нужно производить дальнейших переключений.
Интересно рассмотреть, что получится при изменении положения пробивок в "хромосоме". При очень небольших изменениях "организм" может получать от "хромосомы" любой набор из трех команд. Одна "хромосома" была перфорирована так, что она подавала стрелкам команды в обратном порядке, т.е. "организм" собирался на ветке в обратном порядке. Естественно, что такая "мутация" совершенно нежизненна, так как при этом не произойдет "воспроизведения". Можно считать, что тележка, снабженная такими обратными командами, имеет мутировавший "ген", выраженный в "фенотипе" следующего "поколения". Такие неправильные приспособления и были установлены на тележке A, которую можно заметить в группе, изображенной на фото III, на четвертой ветке (крайней справа) и на которой надпись "head" ("голова") перевернута, чтобы показать, что "мозг" этой тележки инвертирован.
На рис.7 показана полная схема всех реле, контактов, "хромосом" и стрелок для веток и тележек ВПУ I. Обозначения элементов схемы следующие. На тележке A:
1. "Хромосома" и направляющая Cr с отверстиями, обозначенными A и B. Направляющая на рисунке заземлена.
2. Контакты Cr1 и Cr2, проводящие команды от "хромосомы" к тележке B.
3. Реле "хромосомы" R; механическая связь этого реле с Cr показана пунктирной линией со стрелкой. Реле попеременно заземляет Cr1 и Cr2 и затем окончательно отключает их от земли.
4. Четыре буферных контакта C1-C4, соединяющие тележки A и B.
5. Рычаг A (тележки A) и выступ F.
На тележке B:
1. Пара реле, управляющих стрелками веток, R1 и R2. Реле R1, будучи возбуждено, подает ток на переключатель открывания ветки S1, и стрелка открывает ветку. При возбуждении реле R2 ток поступает на переключатель замыкания ветки S2, и ветка закрывается.
2. Пара контактов "глаз" ЕА, чувствительных к рычагу A (тележки A); пара контактов "глаз" ЕB, чувствительных к рычагу A (тележки B); обе пары "глаз" реагируют на тележки, проходящие по дорожке среды.
3. Пара контактов ЕАВ или "глаз А или В", реагирующий на выступ на тележке, когда она проходит на следующую ветку.
4. Четыре буферных контакта C1-C4.
5. Рычаг А (тележки В) и выступ F.
Кроме того, на обеих тележках стоят двигатели и источники питания, получающие энергию от рельс. Двигатель тележки А выключается, когда тележка доходит до конца ветки, вследствие размыкания цепи от положительного полюса блока питания на полотне ветки. Но двигатель тележки В продолжает работать и движение вызывает давление в контактах, обеспечивающее надежное соединение между тележками.
Последовательность операций, приводящая к сборке организмов, следующая:
1. Оператор, действуя органами управления "стрелки Провидения" на главном пульте управления, собирает "организм" на первой ветке.
2. Буферный контакт C1 заземляет контакты EA через платформочку Cr1.
3. Прохождение тележки A по дорожке среды вызывает замыкание контактов EA под действием рычага A (тележки A).
4. Вследствие этого возбуждается реле открывания ветки R1, и переключатель S1 открывает ветку.
5. Тележка входит на вторую ветку.
6. Когда тележка А проходит стрелку второй ветки, действие выступа F на входящей тележке вызывает замыкание контактов "глаза" EAB.
7. Замыкание контактоз EAB вызывает возбуждение реле закрывания ветки R2, которое закрывает ветку посредством переключателя S2.
8. Одновременно с (7) замыкание контактов EAB вызывает возбуждение реле "хромосомы" R через платформочку C3. "Хромосома" Cr благодаря этому передвигается вперед во второе положение.
9. Вследствие этого один полюс контактов "глаза" EB заземляется через C2 и Cr2.
10. Прохождение тележки B по дорожке среды вызывает замыкание контактов EB под действием рычага A (тележки B).
11. Вследствие этого возбуждается реле открывания ветки R1, заставляющее переключатель S1 открыть ветку.
12. Тележка B входит на вторую ветку.
13. Когда тележка B проходит через стрелку второй ветки, выступом F на тележке B замыкаются контакты "глаза" EAB.
14. Замыкание контактов EAB вызывает возбуждение реле ветки R2, закрывающее ветку посредством переключателя S2.
15. Одновременно с (14) замыкание контактов EAB вызывает возбуждение реле "хромосомы" R через C3. Вследствие этого "хромосома" Cr передвигается в третье положение.
16. При третьем положении "хромосомы" Cr "глаза" EA и EB, а также Cr и Cr2, отключаются от земли, и вследствие этого "организм" на первой ветке больше не "воспринимает" и не "передает" информации.
17. "Организм" на второй ветке повторяет этапы (2-16) по отношению к третьей ветке; процесс продолжается до тех пор, пока все ветки не будут заполнены или не будут исчерпаны все запасы частей или не произойдет какая-нибудь ошибка.
Случайное столкновение тележек A и B на дорожке среды не имеет значения, так как тележки могут действовать, только находясь на ветке, и положение "хромосомы" может измениться только тогда, когда она пропускает тележку на следующую ветку.
Оказалось абсолютно необходимым принятие одной предосторожности, а именно введение "системы задержки" тележек.

Рис.8. Схема системы задержки.
Если тележки будут свободно двигаться вместе по дорожке среды, они могут подходить друг к другу вплотную. В таком случае, когда" первая тележка будет направлена на ветку, вторая тележка вследствие конечной длины стрелочного механизма обязательно проскочит за ней на ветку, прежде чем стрелка будет закрыта. Это приведет к окончанию цикла. Поэтому было придумано устройство, обеспечивающее, чтобы две тележки не подходили слишком близко одна к другой, когда первая тележка подходит к стрелке. Схема этого устройства изображена на рис.8, и некоторые его элементы можно видеть на фото I-III. Оно представляет участок полотна, которой обесточивается, как только тележка сходит с него и входит на находящийся перед ним активный участок. Только достигнув конца активного участка, расположенного перед стрелками, тележка снова подает ток на "мертвый" участок. Следующая тележка, пройдя "мертвый" участок, в свою очередь обесточит его для следующих тележек. Действие "системы задержки" выражается в том, что тележки выходят с участка поодиночке. Передний конец участка расположен непосредственно перед закругленным активным участком, находящимся перед разветвлениями. Таким образом, когда тележка достигает разветвления, следующая тележка подходит за ней, но не вплотную, так как скорости тележек примерно одинаковы. Устройство служило не только для предупреждения неполадок при сборке, но и для того, чтобы тележки не толкали друг друга на закругленном участке и не сходили с рельс. Действие "системы задержки" можно рассмотреть на фото II, на котором видно, что шесть тележек задержаны на "мертвом" участке пути, одна тележка находится на активном участке и одна готова войти на вторую ветку. Две тележки вошли на первую ветку. Ясно видно, что тележки на закругленном участке и в разветвлениях отделены друг от друга, так как накопление тележек происходит на прямом "мертвом" участке.
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Чт Май 20, 2021 1:41 am
ПРИЛОЖЕНИЕ Б. УСТРОЙСТВО МОДЕЛЕЙ ВПУ II-V
Опишем теперь подробности устройства цепей и последовательностей сборки более полных моделей "воспроизведения", обозначенных в тексте статьи ВПУ II-V. Эти модели не были изготовлены. Как видно из их устройства, трудность изготовления этих моделей различна. В принципе все они могут работать.
Сперва опишем самовоспроизводящуюся размножающуюся модель ВПУ II, схема которой приведена на рис.6. На нем видно, что каждый "организм" имеет два "отпрыска". Стрелки указывают, какой "отпрыск" принадлежит "организму", находящемуся на данной ветке. На изображенной системе дорожек можно разместить пятнадцать "организмов", принадлежащих четырем "поколениям".
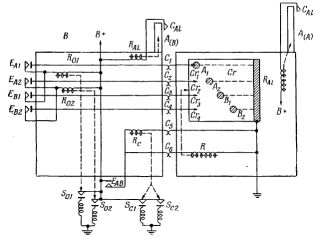
Рис.9. Электрическая схема тележек ВПУ II.
На рис. 9 показана электрическая цепь тележек ВПУ II. Она отличается от цепи тележек ВПУ I следующими особенностями:
1. "Хромосома" Cr имеет четыре отверстия, A1, B1, A2, B2. Эти обозначения указывают, что тележка A находится на первой ветке, тележка B - на первой ветке, тележка A - на второй ветке и тележка B - на второй ветке. При такой схеме ветки "отпрысков" заполняются в порядке A1, A2, B1, B2, хотя, конечно, возможны другие порядки. Первая и вторая ветки на рисунке обозначают две ветки "отпрысков", как указывают стрелки, от ветки "родителей".
2. Имеются соответствующие дополнительные контакты "хромосомы" Cr3 и Cr4 и пружинные контакты C5 и C6.
3. На тележке B имеется вдвое большее число цепей "глаз". Практически нужны лишь два "глаза"; это должны быть перекидные двухполюсные контакты. Имеются также два реле открывания веток "отпрысков" Ro1 и Ro2. Нужно лишь одно реле закрывания ветки, Rc; оно сразу закрывает обе ветки "отпрысков".
За исключением этого увеличения деталей цепи тождественны с цепями ВПУ I. Читатель может легко проследить последовательность сборки, приводящую к заполнению обеих веток "отпрысков".
Когда несколько "организмов" воспроизводятся одновременно, они могут мешать друг другу. Например, рычаг тележки A может открыть две ветки, когда он проходит перед "глазами" A двух воспроизводящихся "организмов". Для предотвращения такой путаницы нужно установить такую последовательность, чтобы одновременно воспроизводил только один "организм". Это удобно сделать следующим образом. Все тележки получают приспособления для опускания рычагов, которые возбуждаются, когда рычаг касается чувствительного, т.е. заземленного "глаза". На рис.9 это выполняет реле RAL. Контакт CAL, касаясь заземленного "глаза", заставляет это реле опустить рычаг A(A) или A(B).

Рис.10. Электрическая схема тележек ВПУ III.
Следующая модель, ВПУ III, не требует стрелок на путях, так как она сама прокладывает для себя ветки со стрелками. Кроме того, в ней должна быть предусмотрена подача тележек на ветку, которой управляют, прямо или косвенно, ранее созданные "организмы". ВПУ III имеет такое же расположение, как ВПУ I, за исключением того, что в ней нет боковых веток. Электрическая схема тележек показана на рис. 10. Она отличается от схемы ВПУ I следующими особенностями.
1. На каждой тележке имеется свой механизм, обозначенный соответственно M(A) и M(B). Механизм M(A), будучи возбужден через CM(A) выполняет в указанном порядке следующие действия: а) останавливает тележку A, б) раскладывает стрелку и ветку перед тележкой A и в) направляет тележку A на ветку. Механизм M(B), будучи возбужден через CM(B) производит только открывание ветки, только что проложенной перед тележкой, посредством тока, посылаемого к выключателю ENS.
2. Каждая тележка имеет контакт, обозначенный CM(A) или CM(B), через который возбуждаются M(A) или M(B), когда соответствующая тележка проходит мимо "организма" на ветке. Их возбуждают через переключатель ENAB реле R1 на тележке A; соответствующее реле открывания ветки R1 на ВПУ I. R1 будучи возбуждено должно оставаться замкнутым достаточное время, чтобы проходящая тележка продвинулась от того места, где ее рычаг замкнул "глаз" на тележке B, до того места, где ее контакт CM коснется переключателя ENAB на тележке B.
3. На каждой тележке имеется рычаг закрывания ветки SC, который автоматически закрывает ветку, когда тележки пройдут через стрелку.
4. В ВПУ III отсутствуют реле закрывания ветки R2, выступы F и "глаз" EAB, имеющиеся в ВПУ I.
Последовательность действий, вполне аналогичная последовательности действий в ВПУ I, следующая:
1. Оператор дает ток в M(A) на тележке А через CM(A). Тележка A останавливается, прокладывает ветку со стрелкой и проходит на ветку. После этого SC закрывает ветку.
2. Затем при приближении тележки B оператор подает ток на M(B). Тогда тележка B подает ток на выключатель ENS, который открывает только что проложенную ветку и впускает тележку В на ветку. После этого SC закрывает ветку. Этим завершается "акт творения". Один контакт "глаза" EA заземляется через C1 и Cr1.
3. При прохождении тележки A по дорожке "среды" рычаг A тележки A ударяет EA и замыкает его.
4. Возбуждается реле R1 на тележке B. При прохождении тележки A реле дает ток в M(A) через выключатель ENAB, который остается под током некоторое время после возбуждения R1.
5. При прохождении тележки A реле M(A) заставляет ее остановиться, проложить вторую ветку со стрелкой и пройти дальше.
6. и 7. Когда другая тележка A входит на вторую ветку, SC закрывает вторую ветку.
8. Одновременно с (5) возбуждается реле R через C3 и C4, и вследствие этого Cr передвигается на второе положение.
9. Один контакт "глаза" EB заземляется через C2 и Cr2.
10. При прохождении тележки B по дорожке "среды" рычаг A тележки B ударяет EB и замыкает его.
11. Возбуждается реле R1 на тележке B. Реле при прохождении тележки B дает ток в M(B) через ENAB.
12. При прохождении тележки B через ENS механизм M(B) открывает вторую ветку.
13. Тележка В проходит на вторую ветку.
14. SC закрывает вторую ветку.
15. Одновременно с (11) возбуждается реле R на тележке А через C3 и C4 заставляет Cr передвинуться на третье ("мертвое") положение.
16. При этом контакты EA и EB отсоединяются от земли, и на первой ветке больше никаких действий не происходит.
17. Организм на второй ветке повторяет этапы (2-16) и т.д.
Очевидно, изготовление ВПУ III связано с большими техническими трудностями. Очень трудно изготовить реле M(A), которое последовательно должно останавливать тележку, проложить довольно большую ветку со стрелкой и направить тележку на проложенную ветку. По-видимому, всего труднее изготовить саму стрелку. Она должна подойти к уже проложенному пути, должна закрывать ветку под действием рычагов SC на тележках A и B, и должно быть предусмотрено, чтобы приближающаяся тележка В с выступающим включенным контактом ENS могла снова открыть ветку. Хотя такой прибор можно признать в принципе осуществимым, для его изготовления потребуется весьма значительное число разнообразных деталей. Стоимость и изготовление такого деликатного, неустойчивого и сложного механизма не оправдывается возможностью практической проверки модели, которая сама прокладывает ветки. Ни одна из описанных моделей ВПУ, кроме ВПУ III, не имеет такого приспособления.
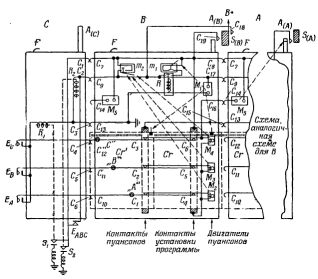
Рис.11. Электрическая схема тележек ВПУ IV.
Опишем теперь модель ВПУ IV - размножающуюся модель, копирующую планы. В ВПУ IV, как описано выше, "организм" превращает пустую карту в "хромосому" с соответствующими пробивками и передает ее тележке, не имеющей "хромосомы". Здесь необходимо "размножающее" перфорирующее устройство и устройство для продвижения и передачи "хромосомы". На рис.3 показано общее расположение модели ВПУ IV. На рис.11 показана электрическая схема тележек, а на рис.12 - последовательность ступеней передвижения "хромосомы" и перфорирования отверстий.
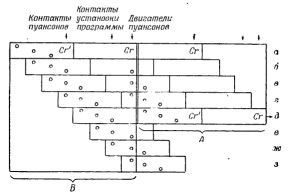
Рис.12. Последовательность передвижения "хромосомы" и воспроизведения в ВПУ IV.
Тележка А имеет следующее оборудование:
1. Пустую карту "хромосомы" Cr, изображенную на рис.11 в нижней левой части тележки A, окруженной пунктиром; на рисунке 12(а) она расположена в левой части тележки.
2. Двигатель "хромосомы" M1. На рис.11 M1 изображен в виде ящика, соединенного с тележкой B. Действие перемещения тележек "хромосомы" вправо указывают две стрелки, ведущие вниз к ящикам Cr и Cr' на тележке B, Стрелки, как видно на рисунке, ведут к двум черным силуэтам маленьких цилиндрических шестерен, перемещающих карты. Карты, изображенные в виде больших пунктирных прямоугольников, снабжены по краям зубчатыми колесиками. Цилиндрические шестерни, зацепляясь с зубчатыми колесиками, перемещают карты. Эти колесики изображаются параллельными, вертикальными прямыми вдоль края карт. Таким образом, нужно представить, что карты под действием двигателя M1 перемещаются горизонтально вправо.
3. Три двигателя для перфорирования "хромосом", M2, M3 и M4. Они изображены в виде маленьких ящиков. Подводки к ним расположены над картой на тележке B. Двигатели перфорируют "хромосомы" в соответствующем положении.
3а. Двигатель изменения рычага M5, который изменяет длину рычага A(A), когда тележка A превращается в тележку B.
4. Реле R, включенное так, что оно само держит свои контакты включенными, и двигатель M1.
5. Миниатюрный прерыватель m1, нормально замкнутый, подающий ток на реле R.
6. Миниатюрный замыкатель цепи m2, нормально разомкнут, подающий ток на реле R.
7. Три контакта, соединяющиеся с пробивками, C1, C2 и C3.
8. Три контакта установки программы, C4, C5 и C6.
9. Набор буферных контактов, C7-C17.
10. Два контакта, подводящие ток от положительного полюса источника питания B к C18 и C19.
11. Выступ F и рычаг A(A).
Тележка B подобна тележке A, но имеет следующие отличия:
1. Она имеет карту "хромосомы" Cr' с пробивками и пустую карту Cr.
2. Рычаг A(B) расположен иначе, чем рычаг A(A).
Тележка С имеет:
1. Реле R1 и R2 для открывания и закрывания веток соответственно через S1 и S2.
2. Контактные "глаза" EA, EB и EC соответственно для распознавания проходящих тележек А, В и С.
3. Контактный "глаз" EABC для распознавания прохождения любой тележки через стрелку следующей ветки.
4. Пружинные буферные контакты C1-C6.
5. Распознающий рычаг A(C) и выступ F.
Ветки имеют два тупиковых упора S(A) и S(B), соединяющиеся соответственно с рычагами A(A) и A(B). Тележки прижимаются друг к другу и к этим двум упорам тяговыми двигателями. Подводка от положительного полюса источника питания B подходит к цепям тележки B от упора S(B) через контакты C18 или C19. Следовательно, все схемы "организмов" реализуются лишь после того, когда тележки собраны на своих ветках, и "жизненный цикл" может начаться лишь после того, как ток от плюса источника питания пройдет в реле R через C18. Он может продолжаться лишь при постоянном соединении с положительным полюсом B через C19. Случайные соединения тележек на дорожке окружения ни к чему не приводят.
На рис.12 изображен подробно цикл перфорирования, перемещения и прохождения карт "хромосомы" (этапы а-з). Пустая карта Cr на тележке B перфорируется аналогично карте Cr'. Затем она передается на тележку A, а пустая карта на тележке A передвигается вперед. После восьми перемещений тележка А имеет такой же комплект карт и пробивок, какой имела тележка В в начале цикла. Вертикальные стрелки на рис.12, так же как на рис.11, указывают положения контактов и двигателей, которые воздействуют на карту или на которые действуют карты в различные моменты цикла.
"Жизненный цикл" протекает следующим образом:
1. Тележки A, B и C собираются по порядку на первой ветке.
2. Вхождение тележки B на ветку подает ток через C18 к первичной обмотке реле R; после этого через короткий промежуток времени, когда C18 проходит упор S(B) и C19 доходит до упора, подается ток через C19. Это приводит к следующим действиям, которые повторяются всякий раз при возбуждении реле R.
Так как контакты реле R нормально замкнуты, ток первичной обмотки реле прерывается срабатыванием прерывателя m1.
Через провода, соединенные параллельно реле R, подается ток в двигатель M1. M1 передвигает обе карты на тележке B, Cr и Cr', пока они не переместятся на 1/8 расстояния между центрами тележек A и B, и тогда размыкается нормально замкнутый прерыватель m1.
Реле R размыкается, и M1 останавливается.
Реле R на тележке A возбуждается одновременно с реле на тележке B током через контакт C17 на тележке B и контакт C9 на тележке А.
На тележке A запускается двигатель M1, который проходит такой же цикл, как двигатель M1 на тележке В, передвигая Cr на тележке A на такое же расстояние, как Cr и Cr' на тележке B.
В этот момент карты "хромосом" переместились в положения, указанные на рис.12(б), но в Сr не пробито никаких отверстий.
3. Когда первое отверстие ("A") доходит до контакта пуансона C1 контакт C1 замыкается. Это приводит к запуску двигателя M2, питание на который подается от положительного полюса B через C19, а соединение с землей производится через отверстие "А" и C1 через C15 на тележке B и C13 на тележке A. Двигатель M2 пробивает первое отверстие в Cr. Это положение изображено на рис.12(б).
4. Двигатель M2 замыкает также переключатель m2, который посылает ток через первичную цепь R и вызывает еще один цикл передвижения "хромосом", как описано в (2).
5. Когда Cr' доходит до такого положения, что отверстие "B" находится против контакта пуансона, замыкается контакт Cr. Это приводит к запуску двигателя M3, и в "хромосоме" M3 делается вторая пробивка, как в (3). Это положение изображено на рис.12(в).
6. После второй пробивки все "хромосомы" передвигаются вперед еще на одну ступень, как в (4).
7. Когда третья пробивка "C" доходит до контакта пуансона, замыкается контакт С3. Это приводит к пуску двигателя пуансона M4, как в (3) и (5). На рис.12(г) изображено положение после того, как сделана третья пробивка.
8. Все "хромосомы" передвигаются еще на одну ступень, как в (4) и (6). Но пуск двигателя M4 приводит к пуску двигателя M5 на тележке A, так как их цепи заземления идут параллельно, через C3 и C15 на тележке B и C13 на тележке A. После некоторой задержки, достаточной для того, чтобы "хромосомы" передвинулись на обеих картах, M5 изменяет длину рычага распознавания A(A) и делает ее такой, как требуется для А(B). Упор S(A) теперь уже не может задерживать тележку A, которая становится тождественной оригиналу тележки В (рис.12(д)), и "новорожденная" тележка В выходит на дорожку среды через возвратную ветку. Вследствие разрыва цепи заземления в C15 на B и C13 на A напряжение не подается на контакты пуансонов, так что их двигатели больше не работают.
9. В этом положении (рис.12(д)), когда новая тележка B отослана на дорожку среды, контакт программы C4 замкнут, и ток от положительного полюса B подводится через C10 на тележке B и C6 на тележке C к одному контакту "глаза" EA, чувствительного к рычагам на тележках A, проходящих по дорожке среды.
10. Когда тележка A проходит EA и от положительного полюса B подается ток на реле R1 на A, от которого цепь заземления идет через C3 на C и C13 на B, R1 открывает при помощи S1 следующую ветку, и тележка A проходит на нее и останавливается упором S(A).
11. Когда тележка A проходит стрелку ветки, "глаз" EABC на первом "организме" замыкается выступом F. Напряжение на EABC все время подается через C1 и C7 на тележке B. При замыкании EABC возбуждается реле R2, закрывающее следующую ветку посредством Sr. Кроме того, Rr дает ток (через контакты C2 на C и C9 на B) реле R на тележке B, которое заставляет передвинуться Cr' на В. Создавшееся положение изображено на рис.12(е).
12. В этом положении контакт программы C5 замкнут и напряжение подается на C13 на тележке С, как в (9).
13. Когда проходит тележка B (имеется только одна тележка В, только что созданная в этом цикле, как описано в (10), вследствие замыкания EB открывается следующая ветка, и тележка входит на эту ветку и останавливается упором S(B). Здесь начинается "жизненный цикл" следующего "организма", так как тележка C участвует в этом цикле только для выполнения программы.
14. После прохождения тележки B на следующую ветку, как описано в (11), EABC и R2 закрывают ветку посредством S2, и "хромосома" на B передвигается в положение, изображенное рис.12(ж).
15. Как описано в (12), замыкается контакт установки программы C6, через который подается ток к контактному "глазу" EC.
16. Как и в (13) тележка C, проходя мимо EC, направляется на следующую ветку для завершения следующего "организма" (который уже начал "размножать" свои планы). Теперь стрелку на эту ветку нужно выключить совсем, поскольку после следующего передвижения "хромосомы" первого "организма" все три контакта C4, C5 и C6 останутся замкнутыми. Для этого можно было бы снабдить тележку C счетчиком, который позволял бы реле R1 до того, как оно "испустит дух", произвести только три действия, но проще применить следующий способ. Последовательно с входной стрелкой ветки устанавливается замыкающий миниатюрный выключатель, который раз мыкается только рычагом A(C) на тележке C. Как только тележка входит на ветку, она открывает этот выключатель, так что реле R1 "организма", находящегося на предыдущей ветке, не сможет опять открыть ветку. Таким образом, на заполненную ветку новые тележки войти не могут.
17. Прохождение тележки C на следующую ветку вызвало замыкание EABC, а R2 закрывает ветку посредством S2. "Хромосома" на В передвигается в положение, изображенное на рис.12(з), и покидает израсходованный "организм". Замыкание программных контактов не оказывает никакого действия, благодаря мерам, описанным в (16). При случайном прохождении тележки мимо "глаз" EA, EB или EC реле R замыкает холостой контакт.
Изготовление модели ВПУ IV ввиду ее сложности должно быть чрезвычайно трудным и дорогим. Без сомнения, понадобятся специальные тележки большого габарита, так как на них должно быть размещено много различных деталей. Значение этого проекта в том, что он доказывает принципиальную возможность изготовления механической модели полного процесса воспроизведения.
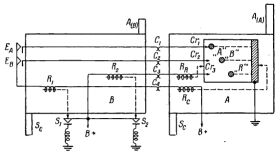
Рис.13. Электрическая схема тележек ВПУ V.
В заключение опишем модель ВПУ V, столь же легко осуществимую, как ВПУ I. ВПУ V представляет устройство, в котором "организмы" "разлагаются после смерти", а их части используются для создания новых "организмов", являющихся воспроизведениями "родителей". Практическая осуществимость этой модели основана на том, что здесь требуется небольшое число тележек, так как старое "поколение" разбирается на части. Схема модели представлена на рис.4. Здесь необходимо не больше четырех тележек, хотя можно использовать и больше. Как только "организм" из двух тележек произвел второй "организм", первый "распадается на части", которые выходят на дорожку среды по возвратной ветке. На рис.13 показана электрическая схема тележек модели ВПУ V. Она имеет следующие отличия от схемы ВПУ I:
1. "Хромосома" имеет третье отверстие, "R" и контакт Cr3, указывающие - "разрушаться и возвращаться в среду".
2. Реле возврата "хромосомы" Rr соединено с Cr3. Будучи возбуждено, оно возвращает "хромосому" Cr в исходное положение, так что она может пойти на дорожку среды и может быть использована снова. Цикличная "хромосома" могла бы устранить необходимость в реле возврата.
3. Реле R2 соединено с Cr3 через контакты C3. Будучи возбуждено, оно открывает упор в конце ветки посредством S2 и пропускает обе тележки обратно на дорожку среды через ветки возврата.
4. Закрытие веток производится автоматически находящимся на тележке рычагом закрывания ветки SС, когда тележка входит на ветку. Это необходимо потому, что реле R2, которое закрывало ветки в ВПУ I, занято открыванием конечного упора ветки.
5. "Хромосома" передвигает реле Rc всякий раз, когда соответствующая тележка закрывает "глаз" EA или EB, так как реле Rc соединено последовательно с реле открывания ветки R1.
Последовательность действий ясна из схемы и сходна с последовательностью действий в ВПУ I с учетом указанных выше различий. Если во всей системе применяются только четыре тележки, "отмершая" пара выходит с боковых веток в том же порядке, как она входила на них, и другому "организму" не нужно проводить никакого упорядочения. Таким образом, это не является настоящей моделью воспроизведения, если не ввести еще одну тележку, чтобы детали на дорожке среды были расположены до некоторой степени случайно.
ЛИТЕРАТУРА
1. von Neumann J., The general and logical theory of automata, in: Cerebral Mechanisms in Bechavior, Wiley, New York, 1951.
2. von Neumann J., Unpublished work; цитируется в статье: Кement J., Man viewed as a machine, Scient. Amer., 192, 4 (1955), 58.
3. Davidson J.N., The Biochemistry of the Nucleic Acids, Methuen, London, 1953, p.192.
4. Jacobson H., The information content of mechanisms and circuits, неопубликованная работа, 1958.
5. Barricelly N.A., The hypothesis of the gene-symbiosis and its importance in the explanation of the origin of the gamic reproduction, Gammermeyers Boghandel, Oslo, 1947.
6. Barricelly N.A., On the manner in which crossbreeding takes place in bacteriophages and bacteria, Ada Biotheoretica, 11, 11 (1955), 75.
7. Haldane J.B.S., The mechanical chess-player, Brit. J. Philos. Sci., 3 (1952), 189.
8. Jacobson H., Information, reproduction, and the origin of life, Amer. Scientist, 43 (1955), 119.
Опишем теперь подробности устройства цепей и последовательностей сборки более полных моделей "воспроизведения", обозначенных в тексте статьи ВПУ II-V. Эти модели не были изготовлены. Как видно из их устройства, трудность изготовления этих моделей различна. В принципе все они могут работать.
Сперва опишем самовоспроизводящуюся размножающуюся модель ВПУ II, схема которой приведена на рис.6. На нем видно, что каждый "организм" имеет два "отпрыска". Стрелки указывают, какой "отпрыск" принадлежит "организму", находящемуся на данной ветке. На изображенной системе дорожек можно разместить пятнадцать "организмов", принадлежащих четырем "поколениям".

Рис.9. Электрическая схема тележек ВПУ II.
На рис. 9 показана электрическая цепь тележек ВПУ II. Она отличается от цепи тележек ВПУ I следующими особенностями:
1. "Хромосома" Cr имеет четыре отверстия, A1, B1, A2, B2. Эти обозначения указывают, что тележка A находится на первой ветке, тележка B - на первой ветке, тележка A - на второй ветке и тележка B - на второй ветке. При такой схеме ветки "отпрысков" заполняются в порядке A1, A2, B1, B2, хотя, конечно, возможны другие порядки. Первая и вторая ветки на рисунке обозначают две ветки "отпрысков", как указывают стрелки, от ветки "родителей".
2. Имеются соответствующие дополнительные контакты "хромосомы" Cr3 и Cr4 и пружинные контакты C5 и C6.
3. На тележке B имеется вдвое большее число цепей "глаз". Практически нужны лишь два "глаза"; это должны быть перекидные двухполюсные контакты. Имеются также два реле открывания веток "отпрысков" Ro1 и Ro2. Нужно лишь одно реле закрывания ветки, Rc; оно сразу закрывает обе ветки "отпрысков".
За исключением этого увеличения деталей цепи тождественны с цепями ВПУ I. Читатель может легко проследить последовательность сборки, приводящую к заполнению обеих веток "отпрысков".
Когда несколько "организмов" воспроизводятся одновременно, они могут мешать друг другу. Например, рычаг тележки A может открыть две ветки, когда он проходит перед "глазами" A двух воспроизводящихся "организмов". Для предотвращения такой путаницы нужно установить такую последовательность, чтобы одновременно воспроизводил только один "организм". Это удобно сделать следующим образом. Все тележки получают приспособления для опускания рычагов, которые возбуждаются, когда рычаг касается чувствительного, т.е. заземленного "глаза". На рис.9 это выполняет реле RAL. Контакт CAL, касаясь заземленного "глаза", заставляет это реле опустить рычаг A(A) или A(B).

Рис.10. Электрическая схема тележек ВПУ III.
Следующая модель, ВПУ III, не требует стрелок на путях, так как она сама прокладывает для себя ветки со стрелками. Кроме того, в ней должна быть предусмотрена подача тележек на ветку, которой управляют, прямо или косвенно, ранее созданные "организмы". ВПУ III имеет такое же расположение, как ВПУ I, за исключением того, что в ней нет боковых веток. Электрическая схема тележек показана на рис. 10. Она отличается от схемы ВПУ I следующими особенностями.
1. На каждой тележке имеется свой механизм, обозначенный соответственно M(A) и M(B). Механизм M(A), будучи возбужден через CM(A) выполняет в указанном порядке следующие действия: а) останавливает тележку A, б) раскладывает стрелку и ветку перед тележкой A и в) направляет тележку A на ветку. Механизм M(B), будучи возбужден через CM(B) производит только открывание ветки, только что проложенной перед тележкой, посредством тока, посылаемого к выключателю ENS.
2. Каждая тележка имеет контакт, обозначенный CM(A) или CM(B), через который возбуждаются M(A) или M(B), когда соответствующая тележка проходит мимо "организма" на ветке. Их возбуждают через переключатель ENAB реле R1 на тележке A; соответствующее реле открывания ветки R1 на ВПУ I. R1 будучи возбуждено должно оставаться замкнутым достаточное время, чтобы проходящая тележка продвинулась от того места, где ее рычаг замкнул "глаз" на тележке B, до того места, где ее контакт CM коснется переключателя ENAB на тележке B.
3. На каждой тележке имеется рычаг закрывания ветки SC, который автоматически закрывает ветку, когда тележки пройдут через стрелку.
4. В ВПУ III отсутствуют реле закрывания ветки R2, выступы F и "глаз" EAB, имеющиеся в ВПУ I.
Последовательность действий, вполне аналогичная последовательности действий в ВПУ I, следующая:
1. Оператор дает ток в M(A) на тележке А через CM(A). Тележка A останавливается, прокладывает ветку со стрелкой и проходит на ветку. После этого SC закрывает ветку.
2. Затем при приближении тележки B оператор подает ток на M(B). Тогда тележка B подает ток на выключатель ENS, который открывает только что проложенную ветку и впускает тележку В на ветку. После этого SC закрывает ветку. Этим завершается "акт творения". Один контакт "глаза" EA заземляется через C1 и Cr1.
3. При прохождении тележки A по дорожке "среды" рычаг A тележки A ударяет EA и замыкает его.
4. Возбуждается реле R1 на тележке B. При прохождении тележки A реле дает ток в M(A) через выключатель ENAB, который остается под током некоторое время после возбуждения R1.
5. При прохождении тележки A реле M(A) заставляет ее остановиться, проложить вторую ветку со стрелкой и пройти дальше.
6. и 7. Когда другая тележка A входит на вторую ветку, SC закрывает вторую ветку.
8. Одновременно с (5) возбуждается реле R через C3 и C4, и вследствие этого Cr передвигается на второе положение.
9. Один контакт "глаза" EB заземляется через C2 и Cr2.
10. При прохождении тележки B по дорожке "среды" рычаг A тележки B ударяет EB и замыкает его.
11. Возбуждается реле R1 на тележке B. Реле при прохождении тележки B дает ток в M(B) через ENAB.
12. При прохождении тележки B через ENS механизм M(B) открывает вторую ветку.
13. Тележка В проходит на вторую ветку.
14. SC закрывает вторую ветку.
15. Одновременно с (11) возбуждается реле R на тележке А через C3 и C4 заставляет Cr передвинуться на третье ("мертвое") положение.
16. При этом контакты EA и EB отсоединяются от земли, и на первой ветке больше никаких действий не происходит.
17. Организм на второй ветке повторяет этапы (2-16) и т.д.
Очевидно, изготовление ВПУ III связано с большими техническими трудностями. Очень трудно изготовить реле M(A), которое последовательно должно останавливать тележку, проложить довольно большую ветку со стрелкой и направить тележку на проложенную ветку. По-видимому, всего труднее изготовить саму стрелку. Она должна подойти к уже проложенному пути, должна закрывать ветку под действием рычагов SC на тележках A и B, и должно быть предусмотрено, чтобы приближающаяся тележка В с выступающим включенным контактом ENS могла снова открыть ветку. Хотя такой прибор можно признать в принципе осуществимым, для его изготовления потребуется весьма значительное число разнообразных деталей. Стоимость и изготовление такого деликатного, неустойчивого и сложного механизма не оправдывается возможностью практической проверки модели, которая сама прокладывает ветки. Ни одна из описанных моделей ВПУ, кроме ВПУ III, не имеет такого приспособления.

Рис.11. Электрическая схема тележек ВПУ IV.
Опишем теперь модель ВПУ IV - размножающуюся модель, копирующую планы. В ВПУ IV, как описано выше, "организм" превращает пустую карту в "хромосому" с соответствующими пробивками и передает ее тележке, не имеющей "хромосомы". Здесь необходимо "размножающее" перфорирующее устройство и устройство для продвижения и передачи "хромосомы". На рис.3 показано общее расположение модели ВПУ IV. На рис.11 показана электрическая схема тележек, а на рис.12 - последовательность ступеней передвижения "хромосомы" и перфорирования отверстий.

Рис.12. Последовательность передвижения "хромосомы" и воспроизведения в ВПУ IV.
Тележка А имеет следующее оборудование:
1. Пустую карту "хромосомы" Cr, изображенную на рис.11 в нижней левой части тележки A, окруженной пунктиром; на рисунке 12(а) она расположена в левой части тележки.
2. Двигатель "хромосомы" M1. На рис.11 M1 изображен в виде ящика, соединенного с тележкой B. Действие перемещения тележек "хромосомы" вправо указывают две стрелки, ведущие вниз к ящикам Cr и Cr' на тележке B, Стрелки, как видно на рисунке, ведут к двум черным силуэтам маленьких цилиндрических шестерен, перемещающих карты. Карты, изображенные в виде больших пунктирных прямоугольников, снабжены по краям зубчатыми колесиками. Цилиндрические шестерни, зацепляясь с зубчатыми колесиками, перемещают карты. Эти колесики изображаются параллельными, вертикальными прямыми вдоль края карт. Таким образом, нужно представить, что карты под действием двигателя M1 перемещаются горизонтально вправо.
3. Три двигателя для перфорирования "хромосом", M2, M3 и M4. Они изображены в виде маленьких ящиков. Подводки к ним расположены над картой на тележке B. Двигатели перфорируют "хромосомы" в соответствующем положении.
3а. Двигатель изменения рычага M5, который изменяет длину рычага A(A), когда тележка A превращается в тележку B.
4. Реле R, включенное так, что оно само держит свои контакты включенными, и двигатель M1.
5. Миниатюрный прерыватель m1, нормально замкнутый, подающий ток на реле R.
6. Миниатюрный замыкатель цепи m2, нормально разомкнут, подающий ток на реле R.
7. Три контакта, соединяющиеся с пробивками, C1, C2 и C3.
8. Три контакта установки программы, C4, C5 и C6.
9. Набор буферных контактов, C7-C17.
10. Два контакта, подводящие ток от положительного полюса источника питания B к C18 и C19.
11. Выступ F и рычаг A(A).
Тележка B подобна тележке A, но имеет следующие отличия:
1. Она имеет карту "хромосомы" Cr' с пробивками и пустую карту Cr.
2. Рычаг A(B) расположен иначе, чем рычаг A(A).
Тележка С имеет:
1. Реле R1 и R2 для открывания и закрывания веток соответственно через S1 и S2.
2. Контактные "глаза" EA, EB и EC соответственно для распознавания проходящих тележек А, В и С.
3. Контактный "глаз" EABC для распознавания прохождения любой тележки через стрелку следующей ветки.
4. Пружинные буферные контакты C1-C6.
5. Распознающий рычаг A(C) и выступ F.
Ветки имеют два тупиковых упора S(A) и S(B), соединяющиеся соответственно с рычагами A(A) и A(B). Тележки прижимаются друг к другу и к этим двум упорам тяговыми двигателями. Подводка от положительного полюса источника питания B подходит к цепям тележки B от упора S(B) через контакты C18 или C19. Следовательно, все схемы "организмов" реализуются лишь после того, когда тележки собраны на своих ветках, и "жизненный цикл" может начаться лишь после того, как ток от плюса источника питания пройдет в реле R через C18. Он может продолжаться лишь при постоянном соединении с положительным полюсом B через C19. Случайные соединения тележек на дорожке окружения ни к чему не приводят.
На рис.12 изображен подробно цикл перфорирования, перемещения и прохождения карт "хромосомы" (этапы а-з). Пустая карта Cr на тележке B перфорируется аналогично карте Cr'. Затем она передается на тележку A, а пустая карта на тележке A передвигается вперед. После восьми перемещений тележка А имеет такой же комплект карт и пробивок, какой имела тележка В в начале цикла. Вертикальные стрелки на рис.12, так же как на рис.11, указывают положения контактов и двигателей, которые воздействуют на карту или на которые действуют карты в различные моменты цикла.
"Жизненный цикл" протекает следующим образом:
1. Тележки A, B и C собираются по порядку на первой ветке.
2. Вхождение тележки B на ветку подает ток через C18 к первичной обмотке реле R; после этого через короткий промежуток времени, когда C18 проходит упор S(B) и C19 доходит до упора, подается ток через C19. Это приводит к следующим действиям, которые повторяются всякий раз при возбуждении реле R.
Так как контакты реле R нормально замкнуты, ток первичной обмотки реле прерывается срабатыванием прерывателя m1.
Через провода, соединенные параллельно реле R, подается ток в двигатель M1. M1 передвигает обе карты на тележке B, Cr и Cr', пока они не переместятся на 1/8 расстояния между центрами тележек A и B, и тогда размыкается нормально замкнутый прерыватель m1.
Реле R размыкается, и M1 останавливается.
Реле R на тележке A возбуждается одновременно с реле на тележке B током через контакт C17 на тележке B и контакт C9 на тележке А.
На тележке A запускается двигатель M1, который проходит такой же цикл, как двигатель M1 на тележке В, передвигая Cr на тележке A на такое же расстояние, как Cr и Cr' на тележке B.
В этот момент карты "хромосом" переместились в положения, указанные на рис.12(б), но в Сr не пробито никаких отверстий.
3. Когда первое отверстие ("A") доходит до контакта пуансона C1 контакт C1 замыкается. Это приводит к запуску двигателя M2, питание на который подается от положительного полюса B через C19, а соединение с землей производится через отверстие "А" и C1 через C15 на тележке B и C13 на тележке A. Двигатель M2 пробивает первое отверстие в Cr. Это положение изображено на рис.12(б).
4. Двигатель M2 замыкает также переключатель m2, который посылает ток через первичную цепь R и вызывает еще один цикл передвижения "хромосом", как описано в (2).
5. Когда Cr' доходит до такого положения, что отверстие "B" находится против контакта пуансона, замыкается контакт Cr. Это приводит к запуску двигателя M3, и в "хромосоме" M3 делается вторая пробивка, как в (3). Это положение изображено на рис.12(в).
6. После второй пробивки все "хромосомы" передвигаются вперед еще на одну ступень, как в (4).
7. Когда третья пробивка "C" доходит до контакта пуансона, замыкается контакт С3. Это приводит к пуску двигателя пуансона M4, как в (3) и (5). На рис.12(г) изображено положение после того, как сделана третья пробивка.
8. Все "хромосомы" передвигаются еще на одну ступень, как в (4) и (6). Но пуск двигателя M4 приводит к пуску двигателя M5 на тележке A, так как их цепи заземления идут параллельно, через C3 и C15 на тележке B и C13 на тележке A. После некоторой задержки, достаточной для того, чтобы "хромосомы" передвинулись на обеих картах, M5 изменяет длину рычага распознавания A(A) и делает ее такой, как требуется для А(B). Упор S(A) теперь уже не может задерживать тележку A, которая становится тождественной оригиналу тележки В (рис.12(д)), и "новорожденная" тележка В выходит на дорожку среды через возвратную ветку. Вследствие разрыва цепи заземления в C15 на B и C13 на A напряжение не подается на контакты пуансонов, так что их двигатели больше не работают.
9. В этом положении (рис.12(д)), когда новая тележка B отослана на дорожку среды, контакт программы C4 замкнут, и ток от положительного полюса B подводится через C10 на тележке B и C6 на тележке C к одному контакту "глаза" EA, чувствительного к рычагам на тележках A, проходящих по дорожке среды.
10. Когда тележка A проходит EA и от положительного полюса B подается ток на реле R1 на A, от которого цепь заземления идет через C3 на C и C13 на B, R1 открывает при помощи S1 следующую ветку, и тележка A проходит на нее и останавливается упором S(A).
11. Когда тележка A проходит стрелку ветки, "глаз" EABC на первом "организме" замыкается выступом F. Напряжение на EABC все время подается через C1 и C7 на тележке B. При замыкании EABC возбуждается реле R2, закрывающее следующую ветку посредством Sr. Кроме того, Rr дает ток (через контакты C2 на C и C9 на B) реле R на тележке B, которое заставляет передвинуться Cr' на В. Создавшееся положение изображено на рис.12(е).
12. В этом положении контакт программы C5 замкнут и напряжение подается на C13 на тележке С, как в (9).
13. Когда проходит тележка B (имеется только одна тележка В, только что созданная в этом цикле, как описано в (10), вследствие замыкания EB открывается следующая ветка, и тележка входит на эту ветку и останавливается упором S(B). Здесь начинается "жизненный цикл" следующего "организма", так как тележка C участвует в этом цикле только для выполнения программы.
14. После прохождения тележки B на следующую ветку, как описано в (11), EABC и R2 закрывают ветку посредством S2, и "хромосома" на B передвигается в положение, изображенное рис.12(ж).
15. Как описано в (12), замыкается контакт установки программы C6, через который подается ток к контактному "глазу" EC.
16. Как и в (13) тележка C, проходя мимо EC, направляется на следующую ветку для завершения следующего "организма" (который уже начал "размножать" свои планы). Теперь стрелку на эту ветку нужно выключить совсем, поскольку после следующего передвижения "хромосомы" первого "организма" все три контакта C4, C5 и C6 останутся замкнутыми. Для этого можно было бы снабдить тележку C счетчиком, который позволял бы реле R1 до того, как оно "испустит дух", произвести только три действия, но проще применить следующий способ. Последовательно с входной стрелкой ветки устанавливается замыкающий миниатюрный выключатель, который раз мыкается только рычагом A(C) на тележке C. Как только тележка входит на ветку, она открывает этот выключатель, так что реле R1 "организма", находящегося на предыдущей ветке, не сможет опять открыть ветку. Таким образом, на заполненную ветку новые тележки войти не могут.
17. Прохождение тележки C на следующую ветку вызвало замыкание EABC, а R2 закрывает ветку посредством S2. "Хромосома" на В передвигается в положение, изображенное на рис.12(з), и покидает израсходованный "организм". Замыкание программных контактов не оказывает никакого действия, благодаря мерам, описанным в (16). При случайном прохождении тележки мимо "глаз" EA, EB или EC реле R замыкает холостой контакт.
Изготовление модели ВПУ IV ввиду ее сложности должно быть чрезвычайно трудным и дорогим. Без сомнения, понадобятся специальные тележки большого габарита, так как на них должно быть размещено много различных деталей. Значение этого проекта в том, что он доказывает принципиальную возможность изготовления механической модели полного процесса воспроизведения.

Рис.13. Электрическая схема тележек ВПУ V.
В заключение опишем модель ВПУ V, столь же легко осуществимую, как ВПУ I. ВПУ V представляет устройство, в котором "организмы" "разлагаются после смерти", а их части используются для создания новых "организмов", являющихся воспроизведениями "родителей". Практическая осуществимость этой модели основана на том, что здесь требуется небольшое число тележек, так как старое "поколение" разбирается на части. Схема модели представлена на рис.4. Здесь необходимо не больше четырех тележек, хотя можно использовать и больше. Как только "организм" из двух тележек произвел второй "организм", первый "распадается на части", которые выходят на дорожку среды по возвратной ветке. На рис.13 показана электрическая схема тележек модели ВПУ V. Она имеет следующие отличия от схемы ВПУ I:
1. "Хромосома" имеет третье отверстие, "R" и контакт Cr3, указывающие - "разрушаться и возвращаться в среду".
2. Реле возврата "хромосомы" Rr соединено с Cr3. Будучи возбуждено, оно возвращает "хромосому" Cr в исходное положение, так что она может пойти на дорожку среды и может быть использована снова. Цикличная "хромосома" могла бы устранить необходимость в реле возврата.
3. Реле R2 соединено с Cr3 через контакты C3. Будучи возбуждено, оно открывает упор в конце ветки посредством S2 и пропускает обе тележки обратно на дорожку среды через ветки возврата.
4. Закрытие веток производится автоматически находящимся на тележке рычагом закрывания ветки SС, когда тележка входит на ветку. Это необходимо потому, что реле R2, которое закрывало ветки в ВПУ I, занято открыванием конечного упора ветки.
5. "Хромосома" передвигает реле Rc всякий раз, когда соответствующая тележка закрывает "глаз" EA или EB, так как реле Rc соединено последовательно с реле открывания ветки R1.
Последовательность действий ясна из схемы и сходна с последовательностью действий в ВПУ I с учетом указанных выше различий. Если во всей системе применяются только четыре тележки, "отмершая" пара выходит с боковых веток в том же порядке, как она входила на них, и другому "организму" не нужно проводить никакого упорядочения. Таким образом, это не является настоящей моделью воспроизведения, если не ввести еще одну тележку, чтобы детали на дорожке среды были расположены до некоторой степени случайно.
ЛИТЕРАТУРА
1. von Neumann J., The general and logical theory of automata, in: Cerebral Mechanisms in Bechavior, Wiley, New York, 1951.
2. von Neumann J., Unpublished work; цитируется в статье: Кement J., Man viewed as a machine, Scient. Amer., 192, 4 (1955), 58.
3. Davidson J.N., The Biochemistry of the Nucleic Acids, Methuen, London, 1953, p.192.
4. Jacobson H., The information content of mechanisms and circuits, неопубликованная работа, 1958.
5. Barricelly N.A., The hypothesis of the gene-symbiosis and its importance in the explanation of the origin of the gamic reproduction, Gammermeyers Boghandel, Oslo, 1947.
6. Barricelly N.A., On the manner in which crossbreeding takes place in bacteriophages and bacteria, Ada Biotheoretica, 11, 11 (1955), 75.
7. Haldane J.B.S., The mechanical chess-player, Brit. J. Philos. Sci., 3 (1952), 189.
8. Jacobson H., Information, reproduction, and the origin of life, Amer. Scientist, 43 (1955), 119.
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Чт Май 20, 2021 1:47 am
Жуть. Для игры, конечно, тяжеловато. Однако посмотрим внимательно "Теорию самовоспроизводящихся автоматов" Джона фон Неймана - ТЕМА #98, АБЗАЦ #1242. Ту самую самовоспроизводящуюся клеточную машину, которая здесь была упомянута. Какие-то квадраты-органы, соединенные каналами передачи сигналов. Шифраторы-дешифраторы... Лента чтения-записи... ... Дык, это же компьютер! Т.е. для того, чтобы играть в самовоспроизведение организмов-программ нам не надо моделировать паровозики и клеточные машины!
И, вот, первая задачка на воспроизведение - самый коротенький этюд из книги Ч.Уэзерелла.-ТЕМА #117, АБЗАЦ #1949.
А затем, поэкспериментировав с самосозерцанием можно поиграть не только в самовоспроизведение, но и в более интересные биологические модели, о которых, например, писал Ричард Докинз в своем "Эгоистичном гене" -ТЕМА #25, АБЗАЦ #1303.
ТЕМА #98, АБЗАЦ #1242. Ту самую самовоспроизводящуюся клеточную машину, которая здесь была упомянута. Какие-то квадраты-органы, соединенные каналами передачи сигналов. Шифраторы-дешифраторы... Лента чтения-записи... ... Дык, это же компьютер! Т.е. для того, чтобы играть в самовоспроизведение организмов-программ нам не надо моделировать паровозики и клеточные машины!И, вот, первая задачка на воспроизведение - самый коротенький этюд из книги Ч.Уэзерелла.-
ТЕМА #117, АБЗАЦ #1949.А затем, поэкспериментировав с самосозерцанием можно поиграть не только в самовоспроизведение, но и в более интересные биологические модели, о которых, например, писал Ричард Докинз в своем "Эгоистичном гене" -
ТЕМА #25, АБЗАЦ #1303.
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Пт Май 21, 2021 12:47 am
Из книги Кораблики и солдатики Олега Орлова и Рюрика Попова.
Сухопутная баталия или игра в солдатики.

В Стране оловянных солдатиков. [Явный плагиат с Уэллса].
О СОЛДАТИКАХ
[Точнее, о коллекционерах].
Кто придумал солдатиков или игру в солдатиков - этого сказать совершенно невозможно.
Глиняных, костяных, деревянных, металлических солдатиков для детских игр ученые-археологи находят в раскопках древних римских и греческих городов и даже в египетских пирамидах. А это значит, что солдатикам тысячи и тысячи лет...
Наверное, с тех пор как люди начали воевать и появились воины и войска, мастера стали делать и игрушечных воинов.
В солдатиков - конных воинов - играли дети скифских племен.
В солдатиков - рыцарей - играли дети средневековья.
В солдатиков Преображенского
и Семеновского полков времен Петра Первого играл мальчик Александр Суворов - будущий полководец.
Рассказывают, что французский король Людовик Четырнадцатый заплатил огромную сумму, 50 тысяч серебряных талеров, за целую армию серебряных солдатиков, которых он купил для своего сына.
Сейчас в мире есть музеи оловянных солдатиков. А во многих военных и художественных музеях хранятся крупные коллекции солдатиков.
Так, в ленинградском Эрмитаже сберегается большая коллекция солдатиков художника и искусствоведа Александра Бенуа.
Собирание коллекций и игра в солдатиков привели к тому, что фирмы, изготовляющие солдатиков, стали выпускать ие только фигурки военных, ио и мирных жителей. Чтобы можно было играть не только в войну, но и заселять оловянным народом игрушечные города, улнцы, площади, дома, даже рынки с ярмарками и цыганскими таборами.
Были наборы - да их и сейчас выпускают - героев известных литературных произведений: Гулливера и лилипутов, иегров из "Хижииы дяди Тома", Робинзона Крузо, Пятницу и дикарей, персонажей из сказок братьев Гримм... И просто индейцев, викингов, рыцарей, пиратов.
Впрочем, только ли для детских игр служат солдатики?

Жил в городе Ленинграде военный человек, Михаил Викторович Люшковский. Родился он еще в прошлом веке. Участвовал в первой мировой войне 1914 года, потом в гражданской. Сражался с фашистами в Великую Отечественную, в звании полковника встретил День Победы. Потом преподавал в военном училище историю военного искусства...
И всю жизиь собирал солдатиков! Всех эпох, всех родов войск!
Каких солдатиков ие мог купить или обменять у других коллекционеров, Люшковский отливал из олова сам.
Было у него солдатиков - пятьдесят тысяч! Если бы выстроить это оловяииое войско в одну шеренгу, шеренга протянулась бы по всему Невскому проспекту от Московского вокзала до Адмиралтейства!
Когда начались съемки фильма "Война и мир", понадобилось точно узнать: как были одеты и вооружены солдаты армии Наполеона и русские солдаты Кутузова.
Вспомнили о знаменитом коллекционере. И Михаил Викторович привез на съемки чемодан с солдатиками. И создателям фильма стало понятно, какие, скажем, были шапки у гвардейцев французского императора,- а были они высоченными, из медвежьего меха... Или - какими были артиллерийские запряжки у пушек, оборонявших от французов Шевардинский редут?
У Михаила Викторовича все нашлось.
Помогал Люшковский снимать и сцены баталий. Он сначала расставлял полки и дивизии солдатиков именно так, как они стояли перед битвой при Бородино, а потом показывал: как двигались в ходе сражения, куда отступали или наступали. А режиссер и его помощники смотрели, записывали, зарисовывали в свои блокноты и потом уже командовали своими армиями - тысячами статистов, одетых в формы войск 1812 года...
Так что когда будешь смотреть фильм "Война и мир", снятый по роману Льва Толстого, вспомни эту небольшую историю.
...Свою коллекцию Люшковский завещал Музею Суворова в Ленинграде, где она и хранится.
КАК ИГРАТЬ В СОЛДАТИКОВ
ПОЛЕ БОЯ И ВОЙСКО
Полем боя для игры в солдатиков может служить стол или большой широкий подоконник. Но можно развернуть войска и на полу. Книги, коробки, бруски дерева могут изображать возвышенности, пересеченную местность. Если солдатиков не очень много, удобнее играть на столе.
Для игры не обязательно соблюдать историческую точность. Ведь любым солдатиком в игре действуют по одним и тем же правилам, будь то индеец или наполеоновский гренадер. Историческая точность важна в том случае, если собираешь коллекцию и изучаешь подлинное сражение, бывшее на самом деле.
В сражении с каждой стороны участвуют три рода войск: пехота, кавалерия и артиллерия. Это армия. Но число солдатиков и пушек, участвующих в сражениях, не постоянно. В одной игре может быть армия, в которой много пехоты и конницы, но нет артиллерии, в другой может участвовать пехота и артиллерия, но без кавалерии.
Можно перед сражением поделить солдатиков на две армии так, чтобы было поровну. А если партнер пришел со своим войском, то можно и "все твои против всех моих". И если армии не одинаковые, то тому, у кого меньше войска, придется крепко подумать, как не дать себя разбить более сильному противнику.
Но все-таки лучше партнерам иметь равные по силе армии.
ХОД ИГРЫ
Играющие ходят по очереди. Сначала один передвигает все свои фигурки, потом стреляет. А после него и в той же последовательности это делает противник.
За один свой ход одна из воюющих сторон может передвинуть всю армию и использовать право выстрела. Потом начинает движение другая армия. И тоже стреляет. Можно солдатиков и технику не двигать, а только стрелять.
В случае если какие-то фигурки не передвинуты, а выстрелы за них произведены, двигать их уже нельзя, так как порядок - сначала движение, а потом стрельба - нарушать не положено.
ПЕРЕДВИЖЕНИЕ СОЛДАТИКОВ
Передвижение происходит по определенной мерке. Мы ее называем делением. Делением может служить палочка или обыкновенная спичка.
Пеший солдатик за один ход передвигается на одно деление.
Конный - на три деления.
Если поле для игры не очень просторное, то можно передвигать пеших и конных на меньшее расстояние - пешего на полделения, конного на одно-два. Как договоритесь сразу.
Конные запряжки и повозки двигаются на два деления.
Грузовики, поезда - на пять.
Легковые машины - на шесть.
Пушку можно везти только тремя солдатиками, запряжкой лошадей или любым транспортом. Тогда пушка продвигается так же, как этот транспорт.
Но сдвигать пушку на одно деление могут два пеших артиллериста. Например, они могут выдвинуть пушку из-за угла укрытия, чтобы стрелять.

СТРЕЛЬБА
Стрелять имеет право любой пеший солдатик, если даже у него и нет в руках ружья - таково правило.
Конные не стреляют, они сражаются врукопашную, а о правилах рукопашной ты прочитаешь дальше.
Чтобы сделать выстрел, нужно поднести к плечу солдатика трубочку. Это как бы ружье солдатика. А в трубочку вкладывается небольшой гвоздик - пуля для ружья.
Гвоздик щелчком пальца посылается в неприятельского солдатика.
Если тот упал после попадания, значит, считается убитым. Если солдатик, упав, роняет и других, то и они считаются убитыми этим же выстрелом.
Когда в сражении участвует много солдатиков, то упавших (убитых) нужно убирать с поля боя.
Пушки тоже стреляют гвоздиком. Но их гвоздик-снаряд должен быть потяжелее и подлиннее. Он вкладывается в ствол пушки.
Если гвоздик-снаряд из пушки попадает в грузовик или в другую технику и солдатики падают от сотрясения, то убитыми считаются только упавшие от прямого попадания и при падении уронившие других. Упавшие от сотрясения в грузовике не считаются убитыми.
Вражеская пушка считается сбитой, если она от попадания опрокинулась или отскочила на одно деление или больше. И не обязательно с одного попадания. По ней могут стрелять несколько пушек. И тогда соединенными усилиями, но только в течение одного хода, они могут сбить пушку.
Если солдатик покачнулся, но не упал (прислонился к стене домика или к пушке и тому подобное), он остается в строю.
Если же ты в свой ход, выстрелив всеми пушками, не сбил пушку противника, то она считается уцелевшей и в следующий ход тебе придется все начинать сначала. Приплюсовать результаты прежнего хода нельзя.
Из пушки можно стрелять только тогда, когда возле нее находятся хотя бы два (не меньше) артиллериста. Они могут пушку заряжать, поворачивать в разные стороны. Но не катить! Для этого должно быть не меньше трех солдат возле пушки.
Если противник намерен захватить пушку, а спасти ее - то есть увезти - нельзя, то один солдат может пушку взорвать. Для этого он в свой ход не стреляет в противника, а объявляет пушку взорванной. И сам тоже ходить в этот ход не может. Нельзя взорвать пушку, около которой никого нет. Но противник, подойдя к пушке, может захватить ее.
Стрелять "в упор" в противника не разрешается. Для выстрела расстояние должно быть не короче длины гвоздика-снаряда.
Никакие резинки или пружинки к пушкам приделывать не полагается. Стрелять можно только щелчком пальца.

РУКОПАШНАЯ
Когда противники сходятся на расстояние, меньше длины гвоздика-снаряда, считается, что началась рукопашная схватка.
Как она происходит?
В старинной игре "в зернь" бросали костяные кубики. На боках этих кубиков были нанесены "зерна" - черные точки. Теперь такие кубики делают из дерева или пластмассы.
Кубики-кости нужны и для нашей игры. Кубик бросают, когда солдатики сошлись врукопашную.
Кубик бросают по очереди.
У кого выпало большее количество очков, тот победил.
Побежденный солдатик снимается с поля боя.
При игре сражается врукопашную не одна пара солдатиков, а обычно несколько пар. Кость бросать надо за каждую пару отдельно.
Иногда сражаются врукопашную не пары, а два солдатика против одного или троих. Тогда за одного солдатика кость бросается один раз, а за двоих (или троих) два раза (или три), а результат складывается. У двух и трех солдатиков шансов победить, конечно, больше.
Но бывает и так: за одного солдатика выброшено 6 очков, а, скажем, за трех - 5 очков (2+2+1) - Значит, один победил троих.
Если выпало одинаковое количество очков обоим противникам, значит, в этот ход не одолел никто и рукопашная может быть продолжена в следующий ход.
Но тот, чей ход будет на очереди, может и не продолжать схватку, а увести своего солдата или подвести на помощь еще несколько.
За конного кидают кость два раза и складывают результаты. В рукопашной, таким образом, конный вдвое сильнее пешего.
Конному легче преследовать отступающих пеших - двигается-то он быстрее пеших.
Если рукопашная уже началась и первые кости брошены, то стрелять больше нельзя и те солдатики, за которых забыли пострелять, в этот ход остаются без выстрелов.
Как правило, стрельбой и рукопашной кончается схватка в данный ход.
Но иногда возникает положение, когда можно противника взять в плен.

ПЛЕНЕНИЕ
Больше шести очков один пеший солдатик выбросить не может за один раз - это самое большое число очков на кубике. Поэтому если на одного пешего напало семь пеших, то в рукопашной нет смысла.
Ведь даже если у каждого из семи выпадет по одному очку, то в сумме у нападающих наберется семь очков.
Исход схватки ясен заранее.
В такой ситуации противника берут в плен.
Семеро солдатиков могут брать в плен одного.
Все пленные солдатики до конца сражения участия в нем не принимают.
После окончания "войны" их возвращают.

КОГДА ИГРА СЧИТАЕТСЯ ВЫИГРАННОЙ
Игра прекращается, когда у одного из играющих не останется достаточного количества солдатиков и пушек, чтобы продолжать сражаться - биться ли врукопашную или стрелять. Это и будет конец игры.

Сухопутная баталия или игра в солдатики.

В Стране оловянных солдатиков. [Явный плагиат с Уэллса].
О СОЛДАТИКАХ
[Точнее, о коллекционерах].
Кто придумал солдатиков или игру в солдатиков - этого сказать совершенно невозможно.
Глиняных, костяных, деревянных, металлических солдатиков для детских игр ученые-археологи находят в раскопках древних римских и греческих городов и даже в египетских пирамидах. А это значит, что солдатикам тысячи и тысячи лет...
Наверное, с тех пор как люди начали воевать и появились воины и войска, мастера стали делать и игрушечных воинов.
В солдатиков - конных воинов - играли дети скифских племен.
В солдатиков - рыцарей - играли дети средневековья.
В солдатиков Преображенского
и Семеновского полков времен Петра Первого играл мальчик Александр Суворов - будущий полководец.
Рассказывают, что французский король Людовик Четырнадцатый заплатил огромную сумму, 50 тысяч серебряных талеров, за целую армию серебряных солдатиков, которых он купил для своего сына.
Сейчас в мире есть музеи оловянных солдатиков. А во многих военных и художественных музеях хранятся крупные коллекции солдатиков.
Так, в ленинградском Эрмитаже сберегается большая коллекция солдатиков художника и искусствоведа Александра Бенуа.
Собирание коллекций и игра в солдатиков привели к тому, что фирмы, изготовляющие солдатиков, стали выпускать ие только фигурки военных, ио и мирных жителей. Чтобы можно было играть не только в войну, но и заселять оловянным народом игрушечные города, улнцы, площади, дома, даже рынки с ярмарками и цыганскими таборами.
Были наборы - да их и сейчас выпускают - героев известных литературных произведений: Гулливера и лилипутов, иегров из "Хижииы дяди Тома", Робинзона Крузо, Пятницу и дикарей, персонажей из сказок братьев Гримм... И просто индейцев, викингов, рыцарей, пиратов.
Впрочем, только ли для детских игр служат солдатики?

Жил в городе Ленинграде военный человек, Михаил Викторович Люшковский. Родился он еще в прошлом веке. Участвовал в первой мировой войне 1914 года, потом в гражданской. Сражался с фашистами в Великую Отечественную, в звании полковника встретил День Победы. Потом преподавал в военном училище историю военного искусства...
И всю жизиь собирал солдатиков! Всех эпох, всех родов войск!
Каких солдатиков ие мог купить или обменять у других коллекционеров, Люшковский отливал из олова сам.
Было у него солдатиков - пятьдесят тысяч! Если бы выстроить это оловяииое войско в одну шеренгу, шеренга протянулась бы по всему Невскому проспекту от Московского вокзала до Адмиралтейства!
Когда начались съемки фильма "Война и мир", понадобилось точно узнать: как были одеты и вооружены солдаты армии Наполеона и русские солдаты Кутузова.
Вспомнили о знаменитом коллекционере. И Михаил Викторович привез на съемки чемодан с солдатиками. И создателям фильма стало понятно, какие, скажем, были шапки у гвардейцев французского императора,- а были они высоченными, из медвежьего меха... Или - какими были артиллерийские запряжки у пушек, оборонявших от французов Шевардинский редут?
У Михаила Викторовича все нашлось.
Помогал Люшковский снимать и сцены баталий. Он сначала расставлял полки и дивизии солдатиков именно так, как они стояли перед битвой при Бородино, а потом показывал: как двигались в ходе сражения, куда отступали или наступали. А режиссер и его помощники смотрели, записывали, зарисовывали в свои блокноты и потом уже командовали своими армиями - тысячами статистов, одетых в формы войск 1812 года...
Так что когда будешь смотреть фильм "Война и мир", снятый по роману Льва Толстого, вспомни эту небольшую историю.
...Свою коллекцию Люшковский завещал Музею Суворова в Ленинграде, где она и хранится.
КАК ИГРАТЬ В СОЛДАТИКОВ
ПОЛЕ БОЯ И ВОЙСКО
Полем боя для игры в солдатиков может служить стол или большой широкий подоконник. Но можно развернуть войска и на полу. Книги, коробки, бруски дерева могут изображать возвышенности, пересеченную местность. Если солдатиков не очень много, удобнее играть на столе.
Для игры не обязательно соблюдать историческую точность. Ведь любым солдатиком в игре действуют по одним и тем же правилам, будь то индеец или наполеоновский гренадер. Историческая точность важна в том случае, если собираешь коллекцию и изучаешь подлинное сражение, бывшее на самом деле.
В сражении с каждой стороны участвуют три рода войск: пехота, кавалерия и артиллерия. Это армия. Но число солдатиков и пушек, участвующих в сражениях, не постоянно. В одной игре может быть армия, в которой много пехоты и конницы, но нет артиллерии, в другой может участвовать пехота и артиллерия, но без кавалерии.
Можно перед сражением поделить солдатиков на две армии так, чтобы было поровну. А если партнер пришел со своим войском, то можно и "все твои против всех моих". И если армии не одинаковые, то тому, у кого меньше войска, придется крепко подумать, как не дать себя разбить более сильному противнику.
Но все-таки лучше партнерам иметь равные по силе армии.

ХОД ИГРЫ
Играющие ходят по очереди. Сначала один передвигает все свои фигурки, потом стреляет. А после него и в той же последовательности это делает противник.
За один свой ход одна из воюющих сторон может передвинуть всю армию и использовать право выстрела. Потом начинает движение другая армия. И тоже стреляет. Можно солдатиков и технику не двигать, а только стрелять.
В случае если какие-то фигурки не передвинуты, а выстрелы за них произведены, двигать их уже нельзя, так как порядок - сначала движение, а потом стрельба - нарушать не положено.
ПЕРЕДВИЖЕНИЕ СОЛДАТИКОВ
Передвижение происходит по определенной мерке. Мы ее называем делением. Делением может служить палочка или обыкновенная спичка.
Пеший солдатик за один ход передвигается на одно деление.
Конный - на три деления.
Если поле для игры не очень просторное, то можно передвигать пеших и конных на меньшее расстояние - пешего на полделения, конного на одно-два. Как договоритесь сразу.
Конные запряжки и повозки двигаются на два деления.
Грузовики, поезда - на пять.
Легковые машины - на шесть.
Пушку можно везти только тремя солдатиками, запряжкой лошадей или любым транспортом. Тогда пушка продвигается так же, как этот транспорт.
Но сдвигать пушку на одно деление могут два пеших артиллериста. Например, они могут выдвинуть пушку из-за угла укрытия, чтобы стрелять.

СТРЕЛЬБА
Стрелять имеет право любой пеший солдатик, если даже у него и нет в руках ружья - таково правило.
Конные не стреляют, они сражаются врукопашную, а о правилах рукопашной ты прочитаешь дальше.
Чтобы сделать выстрел, нужно поднести к плечу солдатика трубочку. Это как бы ружье солдатика. А в трубочку вкладывается небольшой гвоздик - пуля для ружья.
Гвоздик щелчком пальца посылается в неприятельского солдатика.
Если тот упал после попадания, значит, считается убитым. Если солдатик, упав, роняет и других, то и они считаются убитыми этим же выстрелом.
Когда в сражении участвует много солдатиков, то упавших (убитых) нужно убирать с поля боя.
Пушки тоже стреляют гвоздиком. Но их гвоздик-снаряд должен быть потяжелее и подлиннее. Он вкладывается в ствол пушки.
Если гвоздик-снаряд из пушки попадает в грузовик или в другую технику и солдатики падают от сотрясения, то убитыми считаются только упавшие от прямого попадания и при падении уронившие других. Упавшие от сотрясения в грузовике не считаются убитыми.
Вражеская пушка считается сбитой, если она от попадания опрокинулась или отскочила на одно деление или больше. И не обязательно с одного попадания. По ней могут стрелять несколько пушек. И тогда соединенными усилиями, но только в течение одного хода, они могут сбить пушку.
Если солдатик покачнулся, но не упал (прислонился к стене домика или к пушке и тому подобное), он остается в строю.
Если же ты в свой ход, выстрелив всеми пушками, не сбил пушку противника, то она считается уцелевшей и в следующий ход тебе придется все начинать сначала. Приплюсовать результаты прежнего хода нельзя.
Из пушки можно стрелять только тогда, когда возле нее находятся хотя бы два (не меньше) артиллериста. Они могут пушку заряжать, поворачивать в разные стороны. Но не катить! Для этого должно быть не меньше трех солдат возле пушки.
Если противник намерен захватить пушку, а спасти ее - то есть увезти - нельзя, то один солдат может пушку взорвать. Для этого он в свой ход не стреляет в противника, а объявляет пушку взорванной. И сам тоже ходить в этот ход не может. Нельзя взорвать пушку, около которой никого нет. Но противник, подойдя к пушке, может захватить ее.
Стрелять "в упор" в противника не разрешается. Для выстрела расстояние должно быть не короче длины гвоздика-снаряда.
Никакие резинки или пружинки к пушкам приделывать не полагается. Стрелять можно только щелчком пальца.

РУКОПАШНАЯ
Когда противники сходятся на расстояние, меньше длины гвоздика-снаряда, считается, что началась рукопашная схватка.
Как она происходит?
В старинной игре "в зернь" бросали костяные кубики. На боках этих кубиков были нанесены "зерна" - черные точки. Теперь такие кубики делают из дерева или пластмассы.
Кубики-кости нужны и для нашей игры. Кубик бросают, когда солдатики сошлись врукопашную.
Кубик бросают по очереди.
У кого выпало большее количество очков, тот победил.
Побежденный солдатик снимается с поля боя.
При игре сражается врукопашную не одна пара солдатиков, а обычно несколько пар. Кость бросать надо за каждую пару отдельно.
Иногда сражаются врукопашную не пары, а два солдатика против одного или троих. Тогда за одного солдатика кость бросается один раз, а за двоих (или троих) два раза (или три), а результат складывается. У двух и трех солдатиков шансов победить, конечно, больше.
Но бывает и так: за одного солдатика выброшено 6 очков, а, скажем, за трех - 5 очков (2+2+1) - Значит, один победил троих.
Если выпало одинаковое количество очков обоим противникам, значит, в этот ход не одолел никто и рукопашная может быть продолжена в следующий ход.
Но тот, чей ход будет на очереди, может и не продолжать схватку, а увести своего солдата или подвести на помощь еще несколько.
За конного кидают кость два раза и складывают результаты. В рукопашной, таким образом, конный вдвое сильнее пешего.
Конному легче преследовать отступающих пеших - двигается-то он быстрее пеших.
Если рукопашная уже началась и первые кости брошены, то стрелять больше нельзя и те солдатики, за которых забыли пострелять, в этот ход остаются без выстрелов.
Как правило, стрельбой и рукопашной кончается схватка в данный ход.
Но иногда возникает положение, когда можно противника взять в плен.

ПЛЕНЕНИЕ
Больше шести очков один пеший солдатик выбросить не может за один раз - это самое большое число очков на кубике. Поэтому если на одного пешего напало семь пеших, то в рукопашной нет смысла.
Ведь даже если у каждого из семи выпадет по одному очку, то в сумме у нападающих наберется семь очков.
Исход схватки ясен заранее.
В такой ситуации противника берут в плен.
Семеро солдатиков могут брать в плен одного.
Все пленные солдатики до конца сражения участия в нем не принимают.
После окончания "войны" их возвращают.

КОГДА ИГРА СЧИТАЕТСЯ ВЫИГРАННОЙ
Игра прекращается, когда у одного из играющих не останется достаточного количества солдатиков и пушек, чтобы продолжать сражаться - биться ли врукопашную или стрелять. Это и будет конец игры.

Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Пт Май 21, 2021 12:54 am
ОПРЕДЕЛИМСЯ С ТЕХНИЧЕСКИМИ ВЫВОДАМИ
Основные вопросы:
1. Применять ли стреляющие пушки? Возражения два: жалко коллекционных миниатюр, и/или трудно достать хорошие пушки. Но, по-моему, лучше уж играть колобашками и потратить время на изготовление чего-нибудь стреляющего. В советское время выпускался набор "Осада крепости". В комплект входили: стандартный строительный набор (бруски, арки, крыши), солдатики (деревянные цилиндры с нарисованными лицами, портупеями и крепящимися сверху киверами) и деревянные стреляющие пушки. Изготовить нечто подобное, думаю, по силам всем.
2. Разрешить ли стрелять (виртуально) солдатикам? Это, в конечном счете, вопрос масштаба. Если считать каждого солдатика моделью одиночного бойца, то, вроде бы, надо. Но не хочется... Мороки много, а увлекательности это не добавляет. Если и вводить виртуальную стрельбу, то как разновидность спецсвойств (снайпера, пулеметчики, ракетометчики...), не забывая и другие спецсвойства (саперы, огнеметчики, минеры...). Тут уместно говорить не о формулах, а об усложнении сценария конкретной игры.
3. По какой формуле рассчитывать рукопашную схватку? Проще всего по Уэллсу, но у него нет правил для раненых. Можно усложнить его правила, считая всех или часть погибших выигравшей схватку стороны не убитыми, а ранеными. Нужна ли в этих формулах случайность? Только в виде исключения, например, обычные солдатики считаются по единице, а герои и боевые роботы "весят" столько, сколько выпадет на кубике.
4. Имеет ли смысл вводить настолько строгие правила, чтобы можно было проводить турниры? Нет. Игра ориентирована в первую очередь на детей, а они должны экспериментировать и фантазировать, пробуя новые формы и способы игры.

В этот раз играть будем не мы...
С сайта www.gardenwargaming.com .
Основные вопросы:
1. Применять ли стреляющие пушки? Возражения два: жалко коллекционных миниатюр, и/или трудно достать хорошие пушки. Но, по-моему, лучше уж играть колобашками и потратить время на изготовление чего-нибудь стреляющего. В советское время выпускался набор "Осада крепости". В комплект входили: стандартный строительный набор (бруски, арки, крыши), солдатики (деревянные цилиндры с нарисованными лицами, портупеями и крепящимися сверху киверами) и деревянные стреляющие пушки. Изготовить нечто подобное, думаю, по силам всем.
2. Разрешить ли стрелять (виртуально) солдатикам? Это, в конечном счете, вопрос масштаба. Если считать каждого солдатика моделью одиночного бойца, то, вроде бы, надо. Но не хочется... Мороки много, а увлекательности это не добавляет. Если и вводить виртуальную стрельбу, то как разновидность спецсвойств (снайпера, пулеметчики, ракетометчики...), не забывая и другие спецсвойства (саперы, огнеметчики, минеры...). Тут уместно говорить не о формулах, а об усложнении сценария конкретной игры.
3. По какой формуле рассчитывать рукопашную схватку? Проще всего по Уэллсу, но у него нет правил для раненых. Можно усложнить его правила, считая всех или часть погибших выигравшей схватку стороны не убитыми, а ранеными. Нужна ли в этих формулах случайность? Только в виде исключения, например, обычные солдатики считаются по единице, а герои и боевые роботы "весят" столько, сколько выпадет на кубике.
4. Имеет ли смысл вводить настолько строгие правила, чтобы можно было проводить турниры? Нет. Игра ориентирована в первую очередь на детей, а они должны экспериментировать и фантазировать, пробуя новые формы и способы игры.

В этот раз играть будем не мы...
С сайта www.gardenwargaming.com .
Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Re: Уэллс, 1913
автор Gudleifr Чт Май 27, 2021 12:44 am
Через 20 лет после "Корабликов и солдатиков" книга была перезапущена, уже одним Р.Поповым - "История моих солдатиков", 2009. Кораблики он убрал. Добавил семейных воспоминаний и много новых красивых картинок. Новых правил почти не вспомнил. К сожалению, в Сеть книгу пока выложили не целиком.



...


...
Обычно пленных у солдатиков не бывает. Взятых в плен пришлось бы отводить под конвоем в тыл. В тылу - содержать под охраной, ослабляя свой фронт. Все это очень неудобно.
Но иногда можно взять в плен вражеского знаменщика или генерала. Так сказать, для огорчения неприятеля...
Если вражеский знаменщик лежит убитый, то, подведя к нему своего солдатика, ты можешь захватить знамя и унести. Само собой, не отрывая, а вместе со знаменщиком. Захват знамени не сделает твое войско сильнее, но противнику будет обидно, потому что утрата знамени - это позор.

СРАЖЕНИЕ
Если войну объявила одна страна другой, то воюют "все мои против всех твоих". А если просто захотелось посражаться, то фигурок можно взять поровну. Но это не обязательно.

Здесь нарисовано поле боя и две армии, готовые начать сражение. На небольших возвышенностях расположились командующие со своими штабами. В центре - селение. Тот, кто овладеет им, сможет обстреливать неприятеля, укрываясь за домами. Это очень выгодная позиция. Ясно, что каждый из противников постарается ее захватить.
Состав армии неодинаков. У "левых" больше кавалерии, у "правых" - пехоты. А каждый пехотинец - это выстрел. И у "правых" есть пушка. Это сразу делает их заметно сильнее.
У кого лучше положение? У "правых". Их пехота несколько ближе к селению. Она быстрее развернется в цепи для наступления, а пушка огнем будет отсекать пехоту "левых", замедляя ее продвижение.
Ну, а как быть "левым"?
У них есть возможности, о которых "правые" не подозревают. Обрати внимание, читатель, как поставлена пушка "правых". Она хорошо простреливает подступы к селению, но от леса до нее рукой подать. "Левым" как можно скорее надлежит собрать к лесу всю свою кавалерию. Для усиления можно присоединить двух генералов с холма и, в виде исключения, взять у пехоты двух конных полковников. Проскакав сквозь лес, кавалерия "левых" атакует пушку. Это будет отчаянная атака в лоб. Разумеется, кавалерия "правых" бросится на выручку, а прислуга постарается пушку увезти. Но у "левых" тут более чем двойное преимущество. У них будет шанс уничтожить пушку, а если повезет, то и захватить ее и увезти к своим. "Левые" могут направить к лесу часть своей пехоты и обстреливать селение, укрываясь за стволами деревьев.
Пехота "правых" окажется под перекрестным огнем, и удержаться в селении ей будет трудно.
Тут не поможет и пушка. Придется отступать к крайним домам, но тогда под прикрытием передних домов, в селение может ворваться кавалерия "левых" и помочь своей пехоте выбить "правых" из селения. Какой вариант выбрать? Тебе решать. Ты - полководец.




...


...
Обычно пленных у солдатиков не бывает. Взятых в плен пришлось бы отводить под конвоем в тыл. В тылу - содержать под охраной, ослабляя свой фронт. Все это очень неудобно.
Но иногда можно взять в плен вражеского знаменщика или генерала. Так сказать, для огорчения неприятеля...
Если вражеский знаменщик лежит убитый, то, подведя к нему своего солдатика, ты можешь захватить знамя и унести. Само собой, не отрывая, а вместе со знаменщиком. Захват знамени не сделает твое войско сильнее, но противнику будет обидно, потому что утрата знамени - это позор.

СРАЖЕНИЕ
Если войну объявила одна страна другой, то воюют "все мои против всех твоих". А если просто захотелось посражаться, то фигурок можно взять поровну. Но это не обязательно.

Здесь нарисовано поле боя и две армии, готовые начать сражение. На небольших возвышенностях расположились командующие со своими штабами. В центре - селение. Тот, кто овладеет им, сможет обстреливать неприятеля, укрываясь за домами. Это очень выгодная позиция. Ясно, что каждый из противников постарается ее захватить.
Состав армии неодинаков. У "левых" больше кавалерии, у "правых" - пехоты. А каждый пехотинец - это выстрел. И у "правых" есть пушка. Это сразу делает их заметно сильнее.
У кого лучше положение? У "правых". Их пехота несколько ближе к селению. Она быстрее развернется в цепи для наступления, а пушка огнем будет отсекать пехоту "левых", замедляя ее продвижение.
Ну, а как быть "левым"?
У них есть возможности, о которых "правые" не подозревают. Обрати внимание, читатель, как поставлена пушка "правых". Она хорошо простреливает подступы к селению, но от леса до нее рукой подать. "Левым" как можно скорее надлежит собрать к лесу всю свою кавалерию. Для усиления можно присоединить двух генералов с холма и, в виде исключения, взять у пехоты двух конных полковников. Проскакав сквозь лес, кавалерия "левых" атакует пушку. Это будет отчаянная атака в лоб. Разумеется, кавалерия "правых" бросится на выручку, а прислуга постарается пушку увезти. Но у "левых" тут более чем двойное преимущество. У них будет шанс уничтожить пушку, а если повезет, то и захватить ее и увезти к своим. "Левые" могут направить к лесу часть своей пехоты и обстреливать селение, укрываясь за стволами деревьев.
Пехота "правых" окажется под перекрестным огнем, и удержаться в селении ей будет трудно.
Тут не поможет и пушка. Придется отступать к крайним домам, но тогда под прикрытием передних домов, в селение может ворваться кавалерия "левых" и помочь своей пехоте выбить "правых" из селения. Какой вариант выбрать? Тебе решать. Ты - полководец.

Gudleifr- Admin
- Сообщения : 3402
Дата регистрации : 2017-03-29
Страница 2 из 2 • 1, 2
Страница 2 из 2
Права доступа к этому форуму:
Вы не можете отвечать на сообщения